HEMOSTASIA
El proceso mediante el
cual el sangrado se detiene se denomina hemostasia. Su función es mantener la integridad de los vasos sanguíneos y el
contenido que fluye a través de los vasos, es decir la sangre.
La hemostasia también se
encarga de mantener la fluidez de la sangre.

FASES DE LA HEMOSTASIA
Hemostasia primaria ( fase
celular de 3-5 minutos)
el objetivo es formar un
TAPÓN INESTABLE
1 vaso-constricción
2 adhesión y agregación
plaquetaria
Hemostasia secundaria
(fase plasmática de 5-10 minutos)
el objetivo es formar un
TAPÓN ESTABLE
3 coagulación
Hemostasia terciaria (de
48-72 horas)
el objetivo es la DISOLUCIÓN DEL COAGULO
4 fibrinolisis
Las tres fases de la
hemostasia se inician al mismo tiempo
FASE VASCULAR
hemostasia primaria
hemostasia primaria
se presenta a través de tres mecanismos:
1 mecanismo miogeno: es un
reflejo del músculo liso de manera que el vaso es colapsado y la sangre tiene
que circular por otra vía disminuyendo así la hemorragia.

2 mecanismo nervioso:produce el reflejo de la contracción del músculo liso.
3 mecanismo humoral: se
liberan una serie de sustancias debido al estrés tales como: serotonina
noradrenalina, endotelina, adrenalina, tromboxano A2

LA VASO-CONSTRICCIÓN
0 la primera fase del
proceso de coagulación es la vasoconstricción.
1 si no se da la
vasoconstricción no puede darse inicio a la siguiente fase de la coagulación.
2 la arteria femoral posee
un calibre tal que es imposible que se desarrolle la vaso-constricción a pesar
de que posee musculo liso. Es por ello que si este vaso se rompiese no existe
posibilidades de que se forme un coagulo.


3 La vaso-constricción es
un mecanismo importante para la fase inicial de la coagulación. No se presenta
en vasos muy grandes (mayores a un centímetro de diámetro como la arteria femoral o la humeral) ni en muy pequeños. En los vasos pequeños existen maneras
de suplir la vaso-constricción y en los vasos muy grandes es posible compensar
este mecanismo fisiológico con la aplicación de un torniquete.
4 la vaso-constricción es
efectiva solamente en los vasos de mediano calibre mientras que en los vasos de
gran calibre, aunque se presenta no es
efectiva. En los vasos de muy pequeño calibre no se desarrolla porque no existe
músculo liso en ellos, para esta función.
5 un ejemplo de tejido que
posee vasos muy pequeños que no poseen músculo liso (capilares) es la encía.
FASE PLAQUETARIA
6 la segunda etapa de la
coagulación es la fase plaquetaria que aparece después de reducir el tamaño de la herida mediante el
mecanismo de vaso-constricción. La fase plaquetaria tiene como objeto la
formación de un tapón plaquetario que impide la salida de sangre por la herida. Este tapón
llamado tapón plaquetario efectivamente está constituido por plaquetas que
circulan en el torrente sanguíneo y se presentan en el momento en que el
endotelio de los vasos se rompe por alguna lesión.




7 las etapas de la fase
plaquetaria:
-adhesión
-activación
-agregación
8 las plaquetas no son
células, son derivados de células. Son fragmentos de citoplasma de una célula
llamada megacariocito.
9 el tiempo de vida de una
plaqueta es de 8 a 13 días.
10 las plaquetas se
encuentran fundamentalmente en algunos órganos en forma de reserva durante su
tiempo de vida, estos órganos son:
Bazo:33%
Pulmón:25%
Por este motivo más de la
mitad de las plaquetas no está en circulación y solamente lo hacen en
situaciones de estrés. Por ejemplo durante una lesión en algunos de los
incontables tramos de vasos circulatorios del organismo. Debido a esta
condición es posible que los resultados observados en un hemograma varíen con
respecto a otro hecho después de algún tiempo en un mismo paciente.
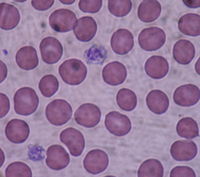
11 las plaquetas tienen
forma redondeada (parecen lentejas) y su numero asciendo entre 150000 y 400000
plaquetas.
12 se caracterizan por
tener en su membrana un fosfolípido especial denominada FACTOR PLAQUETARIO
TRES.
13 también se caracterizan
por presentar en su membrana los receptores de membrana GP. Son varios y vas desde GP1….GP9.
14 las plaquetas vistos al
microscopio presentan gránulos:
-gránulos alfa: que poseen
factores de la coagulación, FACTOR CINCO Y FACTOR OCHO
-gránulos densos: estos
presentan sustancias que activan a otras plaquetas como el ADP, ATP y
serotonina. Por ello estos gránulos están más en relación con la agregación
plaquetaria.
-Gránulos lisosomales
-gránulos de glucógeno: tiene la reserva de energía.
15 las plaquetas como ya
dijimos provienen de los megacariocitos y estos de los megacarioblastos y estos a su vez de unas
células denominadas de la línea mieloide.


16 el megacariocito emite
unas especies de seudópodos que salen por entre las células endoteliales hacia
el torrente sanguíneo de manera que la fuerza del flujo sanguíneo fragmenta
estos seudópodos de forma que se desprenden partes del citoplasma de este
megacariocito debido a la fuerza del flujo sanguíneo. A estos fragmentos les denominamos plaquetas.




17 las plaquetas por lo
tanto no son consideradas células sino fragmentos de células.
18 las plaquetas utilizan
la energía que pudieron acumular del citoplasma del megacariocito antes de
desprenderse. Después del consumo de esta energía las plaquetas se convierten en elementos inertes.
19 el megacariocito es denominado
así porque posee un núcleo muy grande:
-mega: grande
-cario: núcleo
-cito: célula
TROMBOPOYESIS
20 sabemos que cada
megacariocito trombocitogénico genera alrededor de 2000 a 7000 plaquetas.
21 la STEM CELL, que es la célula madre
multi-potencial da origen a la UNIDAD FORMADORA DE COLONIAS S
(CFU-S) y este a su vez de origen a la UNIDAD FORMADORA DE COLONIA
DE MEGACARIOBLASTOS (CFU-MEG) estos dos pasos se dan por mitosis.
22 la CFU-MEG se convierte
en PROMEGA-CAROBLASTO y este después en MEGACARIOBLASTO estos dos procesos se
dan ya no por mitosis sino por endomitosis.
23 la endomitosis es el
proceso por el cual la célula se prepara para dividirse duplicando su material
genético y su citoplasma pero no se divide. Este proceso se puede llegar a
repetir varias veces en el megacarioblasto de manera que se puedan tener desde
8n hasta 32n de material genético. Al final de estas divisiones se genera una
célula muy grande denomina megacariocito.


24 el megacariocito seguidamente forma sus respectivos
gránulos que después pasarán a las plaquetas e inmediatamente empieza a
compartimentalizarse, es decir a la formación de tabiques definiendo así una
estructura que se predispone a fragmentase. Debemos saber que estos fragmentos
están cubiertos de membrana.
25 todo el proceso
anteriormente descrito se denomina trombopoyesis y dura entre 5 y 10 días.
26 la trompoyesis es
dependiente de la trombopoyetina. La característica más importante que se debe
recordar es que la trombopoyetina está presente en todo el proceso de la
trompopoyesis desde la stem cell hasta la célula plenamente funcional que es el
megacariocito.


27 la producción de
trombopoyetina en su mayor medida se realiza en el hígado y en menor proporción
en el riñón.
28 cuando se produce una
herida, es decir la rotura de tejidos, entre ellos: tejido epitelial, tejido
conectivo y dentro de estos los vasos sanguíneos los elementos en salir en
primer lugar son las plaquetas. Esto se debe a que las plaquetas se encuentran
circulando siempre adyacentes al endotelio vascular de manera que cualquier
trauma de este tejido permite inmediatamente la salida de las plaquetas.
Debemos apuntar también que las plaquetas se encuentran por así decirlo en
busca de cualquier desnivel, rugosidad o accidente en el endotelio que en su
mejor estado debe ser liso y continuo.
29 las plaquetas se
desplazan adyacentes a la pared endotelial debido a un fenómeno físico. Las plaquetas quedan relegadas en su mayoría en
las cercanías de la pared endotelial porque en el centro del vaso sanguíneo se
desplazan fundamentalmente los elementos de mayor densidad que son los glóbulos
rojos. Recordemos que las plaquetas miden aproximadamente dos micras mientras
que los glóbulos rojos miden 7 micras. La cantidad de glóbulos rojos es diez
veces mayor que la de plaquetas.


30 al romperse el vaso
sanguíneo las plaquetas atravesaran algunos niveles de estructura. Desde la luz
del endotelio hacia afuera del vaso encontramos el endotelio, después la
membrana basal como parte estructural normal de los tejidos de tipo epitelial en seguida nos encontraremos con una estructura de tejido conjuntivo, que es la
matriz subendotelial compuesto por muchas fibras de colágeno. La plaqueta al
pasar a través de estas estructuras queda aprisionada en las fibras
colágenas. Las plaquetas quedan fijadas en la estructura de fibras colágenas
porque tiene un receptor para estas proteínas que es el GP-Ib, estos recetores
que se encuentran en las plaquetas siempre están activados pero como en la
sangre no hay colágeno entonces las plaquetas siempre están libres.
31 a la unión entre la
plaqueta y el colágeno por intermedio del receptor GP-Ib se le denomina
ADHESIÓN plaquetaria. Por otro lado la unión entre plaquetas se denominaría la
COHESIÓN PLAQUETARIA.
32 la unión entre el
colágeno subendotelial y las plaquetas por medio de los receptores GP-Ib puede ser potenciado por el factor de VON
WILLEBRANSD, aunque no es imprescindible es necesario para que la unión entre plaquetas
y colágeno subendotelial por medio del receptor GP-Ib sea más fuerte.
33 en las enfermedades que
afectan la hemostasia son en su mayoría por una deficiencia en el factor de VON
WILLEBRAND, que como ya dijimos actúa en la fase de ADHESIÓN.
ACTIVACIÓN PLAQUETARIA
34 este proceso se dan los
siguientes procesos:
-metamorfosis viscosa
Cambio de forma
Formación de seudópodos
-activación de GP IIB/IIIA
-síntesis de tromboxano A2
-secreción(liberación) del
contenido de los gránulos de la plaqueta
-activación pro-coagulante
de la membrana (FP-3), es decir la exposición del FECTOR PLAQUETARIO TRES.
35 cuando la plaqueta se
activa se dilata de la mayor forma posible, también emite
seudópodos. La plaqueta por lo tanto intenta cubrir el mayor área posible en la
región del trauma que ha sufrido el vaso sanguíneo. A todo este fenómeno le
denominados metamorfosis viscosa.




36 una vez que la plaqueta
se ha dilatado y ha emitido los seudópodos, es decir se ha llevado a cabo la
metamorfosis viscosa se da la activación de GP IIB/IIIA, que no se encontraban
activadas a diferencia del GP-Ib.




37 después de estos dos
fenómenos la plaqueta libera su contenido, que se encuentra dentro de las
plaquetas. Estos elementos tienen como función activar a las demás plaquetas.
Estos son:
-ADFP
-serotonina
-adrenalina
-tromboxano IIA
-noradrenalina
-endotelina
De esta forma aunque solo algunas plaquetas se activaron al entrar en contacto con el colágeno muchas más plaquetas se activan al ser estimuladas por las plaquetas que liberan su contenido.
De esta forma aunque solo algunas plaquetas se activaron al entrar en contacto con el colágeno muchas más plaquetas se activan al ser estimuladas por las plaquetas que liberan su contenido.
38 si la membrana de una
plaqueta toca la membrana de una plaqueta activada se activa inmediatamente.
Estas plaquetas activadas secundariamente van a liberar sus gránulos.
AGREGACIÓN PLAQUETARIA
39 decimos que las
plaquetas así activadas se van agregar en un proceso que llamamos agregación
plaquetaria.
40 las plaquetas que se
encuentran en fase de agregación plaquetaria van activar sus receptores GP
IIB/IIIA, que son los receptores para fibrinógeno.
41 el fibrinógeno es la
proteína de mayor concentración después de la albumina, en otras palabras
diremos que el fibrinógeno es la que presenta mayor concentración dentro del
grupo de la globulinas.
42 el fibrinógeno es una
proteína en forma fibrilar o de filamento. Esta proteína es simétrica es decir
tiene ambos extremos idénticos. Gracias a esta estructura el fibrinógeno se une a los receptores GP
IIB/IIIA que se encuentran en las plaquetas de esta manera las plaquetas pueden quedar unidas. El fibrinógeno actúa como un puente entre las plaquetas.
43 el proceso
anteriormente descrito se realiza a partir de las primeras plaquetas que quedan
atrapadas en el colágeno sub-endotelial que se activaron para tratar de cubrir
el mayor área posible de la lesión. Después de liberar los gránulos
plaquetarios y activar a las demás plaquetas estas se agregan para formar así
el tapón plaquetario.
44 Gracias a todo este proceso debe ser posible detener el sangrado pero como bien sabemos la hemostasia es un proceso
todavía más largo que continua en una fase siguiente aun. La razón de porque la
hemostasia continua es porque el tapón plaquetario que se ha formado es muy débil
aun, es decir las plaquetas que están unidas a la matriz sub-endotelial aún son
bastante frágiles porque pueden ser arrancadas de este lugar de la lesión por
la fuerza del flujo sanguíneo.
45 la hemostasia primaria
o fase plaquetaria de la hemostasia tiene
como función formar un tapón inestable. Que es todo lo que hemos visto hasta
ahora.
46 a mayor cantidad de AMPc la plaqueta se
mantiene inactiva, es decir se produce la inhibición plaquetaria.
47 a menor cantidad de
AMPc en la plaqueta entonces la plaqueta se activa.


48 Para
que la plaqueta se mantenga con vida esta debe tener AMPc dentro de ella, de
forma que realice los procesos metabólicos que pueda realizar. Cuando la plaqueta
se activa libera el AMPc de forma que su metabolismo cesa y la plaqueta pasa a
ser un elemento inerte. En conclusión
diremos que la activación plaquetaria depende de los niveles de AMPc.
49 EL PRINCIPAL ACTIVADOR
PLAQUETARIO ES EL TROMBOXANO IIA.
50 el tromboxano A2
proviene de los fosfolípidos que están anclados en la parte interna de la membrana de la plaqueta mediante el
siguiente proceso: a través de una enzima denominada fosfolipasa A2 se
liberan ácidos grasos entre ellos el
ácido araquidónico. La enzima ciclo-oxigenasa actúa sobre el ácido graso
mensionado formando los endoperoxidos.
El endoperoxido a través de la tromboxano-sintetasa forma tromboxano A2 que
ejerce una acción inhibitoria sobre la adenilciclasa que es la
responsable de la formación de AMP a partir de ATP. El AMP es el sustrato para
la AMPc que se forma por intervención de la fosfodiesterasa. La formación de troboxano A2
permite la inhibición de AMP cíclico con lo cual la plaqueta pasa a un
estado de activación por disminución de AMPc.
 En la figura de arriba la franja roja representa el endotelio, el fondo blanco la luz de un vaso sanguíneo y el circulo rojo representa la plaqueta.
En la figura de arriba la franja roja representa el endotelio, el fondo blanco la luz de un vaso sanguíneo y el circulo rojo representa la plaqueta.

51 La interacción entre la plaqueta y el endotelio es muy estrecha tanto es así que existe comunicación o señalización celular entre plaqueta y célula endotelial. En principio los endoperoxidos de la plaqueta pueden ingresar a través de la membrana del endotelio vascular y favorecer, a través de la enzima prostaciclina sintetasa, la formación de prostaciclina, la cual ingresa en la plaqueta a través de su membrana y funciona como un activador de la adenilciclasa favoreciendo así la formación de AMPc de forma que la plaqueta se mantiene fuera de el estado de activación por inhibición de adenilciclasa por tromboxano A2. Por lo tanto el endotelio es un regulador de las plaquetas.
52 en base a lo dicho
anteriormente, si se presenta una lesión en el endotelio vascular la plaqueta
no recibirá la prostaciclina de forma que el tromboxano actuará libremente en
la plaqueta activando así a la plaqueta. El endotelio ayuda a evitar que la plaqueta se active. Por esta razón el endotelio es considerado un regulador de la coagulación.
FASE DE LA COAGULACIÓN
53 la coagulación
sanguínea (conocida también como cascada de la coagulación) es parte de la
hemostasia y es una etapa en la que el fibrinógeno se convierte en fibrina.
Esta etapa es posterior a la formación de tapón plaquetario o coagulo
plaquetario temporal.


54 el fibrinógeno es
soluble mientras que la fibrina es una proteína insoluble.
55 los monomeros de
fibrinógeno se unen entre ellos para formar una red de fibrina. Por lo tanto el
monómero es el fibrinógeno mientras que el polímero sería la fibrina.
56 la coagulación es el
paso del cambio de estado de una proteína soluble hacia otra proteína
insoluble, gracias a la intervención de las enzimas que interviene en el
proceso.
57 en las inmediaciones del
tapón plaquetario se agregan los monómeros de fibrinógeno para formar la fibrina, esta última por ser más densa cae sobre el tapón plaquetario de manera que
al caer atrapa glóbulos rojos y convierte en tapón blanco(tapón plaquetario) en
tapón rojo. Por lo tanto el tapón rojo es la unión del papón plaquetario o
blanco más una de red de fibrina insoluble que atrapa en su proceso a glóbulos
rojos y otros elementos.




58 en la práctica haremos
un prueba denominada tiempo de sangría. El procedimiento consiste en producir
una pequeña herida con una lanceta que consiste en observar el sangrado durante un determinando tiempo que es el necesario para que el organismo pueda formar el tapón plaquetario inestable, de
forma que detenga el sangrado. Por lo tanto el tiempo de sangría mide el tiempo
de formación del tapón inestable.


59 Una persona que tenga problemas de hemostasia puede deberse a dos procesos diferentes: 1 relacionada con las plaquetas por lo cual el tiempo de sangría es indefinido o
muy largo, debido a que nunca se forma el tapón plaquetario inestable. 2 O
también puede estar relacionado con la coagulación es decir con la formación de
la fibrina.
60 en los experimentos
veremos que si vertemos sangre en un tubo de ensayo que no tenga un
recubrimiento interior de un anticoagulante(heparina) esta sangre formará un
coagulo distinguiéndose un líquido amarillento y una masa de color rojo.
Después de algún tiempo más este coagulo se encoge debido a que la red de
fibrina se contrae haciendo que el coagulo se reduzca de volumen.
61 la coagulación es una
vía enzimática de múltiples pasos que actúan en cascada. Y se da a través de
dos vías:
-Vía intrínseca
-Vía extrínseca
62 estos dos vías van contribuyen mutuamente en la formación de fibrinógeno.

63 los factores de la coagulación están ordenados por la numeración romana de forma estándar.

63 los factores de la coagulación están ordenados por la numeración romana de forma estándar.

64 los factores de
coagulación son los siguientes:
I: fibrinógeno
II: protrombina
III: factor tisular……………tromboplastina
IV: calcio
V: proacelerina
VII: proconvertina
VIII: F. anti-hemofílico A
IX: F. anti-hemofílico B
X: F. Stuart
XI: factor anti-hemofílico
C
XII:F.Hageman
XIII: factor estabilizador
de la fibrina
64 los factores tienen dos denominaciones activados y desactivados, lo cual quiere decir que pueden estar en dos estados:
64 los factores tienen dos denominaciones activados y desactivados, lo cual quiere decir que pueden estar en dos estados:
-activados: numero romano
simple
-desactivados: numero
romano más la letra “a” en minúscula.
65 una observación de
Perogrullo seria que son doce factores del I al XIII porque el factor numero VI
fue eliminado de manera que solo quedan doce factores.
66 todos los factores de
coagulación son proteínas simógenas a excepción de factor IV que es el calcio
un ión. Sabemos que las proteínas simogenas son aquellas enzimas que se
encuentran desactivadas en su primer estado de existencia. (recordar el
tripsinogeno). Los factores por lo tanto son enzimas que van a participar en la
cascada de coagulación pero que en su primera etapa se encuentran desactivadas
debido a que poseen un fragmento de péptido que le impide activarse. Cuando
este fragmento de péptido se rompe por acción de otra enzima entonces pasa al
estado activado y por lo tanto puede cumplir su función de activar a otro
factor.
67 las concentraciones de
los factores de coagulación son mínimas a excepción del fibrinógeno que puede
ser cuantificado por métodos clínicos se dice que es dosable. Estas enzimas se
encuentran en bajas concentraciones porque su poder catalítico es muy fuerte.
Debemos saber que estas enzimas de la coagulación son parientes de las enzimas
digestivas o enzimas proteolíticas como la quimiotripsina.


68 el proceso de la
coagulación depende de la presencia de los doce factores, de manera que si
uno se encuentra ausente la cascada de coagulación se interrumpe y la
hemostasia no se llega a completar. Por ello decimos que el proceso es muy
delicado y que depende mucho más de los factores que se encuentran en menor
concentración, por lógica. El factor que tenga menor
vida media es el factor crítico en la coagulación y este es el FACTOR VII.
69 todas la proteínas de
la cascada de la coagulación son producidas en el hígado. De manera que si
podemos medir la producción del factor VII estaremos determinando
indirectamente la producción de las demás proteínas. Sabemos que el hígado
produce gran parte de las proteínas que el organismo usa y entre ellas se
encuentra las proteínas de la coagulación de manera que si la producción de
proteínas en el hígado se detiene se verá afectada la
coagulación y dentro de las proteínas de la coagulación que inmediatamente se
ven afectadas mucho más rápido que las demás es el factor VII. Podemos determinar el correcto funcionamiento hepático por
medio de una prueba de coagulación.
70 la coagulación
sanguínea la podemos dividir en tres etapas:
-primera: complejo
pro-trombinasa
-segunda: trombina
-segunda: trombina
-tercera: fibrina
71 EL OBJETIVO DE LA
COAGULACIÓN ES FORMAR LA FIBRINA
72 la fibrina proviene del
fibrinógeno para que esta conversión se dé interviene la trombina. Es decir la
trombina activa al fibrinógeno para convertirlo en fibrina.




73 debemos saber que en el
mismo momento que la trombina activa al fibrinógeno para convertirlo en fibrina
también activa al factor XIII en factor XIIIa es decir el factor estabilizador
de la fibrina en factor estabilizador de la fibrina activado.
74 la TROMBINA es formada por el complejo pro-trombinasa y el complejo pro-trombinasa está formado por
el factor X, el factor V, el calcio y el factor plaquetario número tres.


75 debemos recordar que el
factor plaquetario v y el VIII son co-enzimas, por lo tanto en el caso del
complejo pro-trombinasa la enzima es el factor X mientras que la co-enzima es
el factor V
76 el factor plaquetario
tres se encuentra en las plaquetas como es obvio.
77 recordemos que la
plaqueta está constituida por una membrana celular la cual como es sabido es
una bicapa lipídica constituida por fosfolípidos diversos. En principio cuando
la plaqueta está en reposo, es decir no está activada el factor plaquetario
número tres se encuentra en la capa de fosfolípidos que mira al citoplasma pero
cuando la plaqueta se encuentra activa este factor plaquetario migra hacia la
capa fosfolipídica externa de forma que ahora el factor plaquetario tres se
encuentra en el exterior de la plaqueta lista para formar el complejo
pro-trombinasa. En conclusión cuando la plaqueta se activa esta expone el
factor plaquetario tres.

78 previamente a la exposición
del factor plaquetario tres el factor V y el factor X están libres en el
torrente sanguíneo. Una vez que se expone el factor plaquetario tres estos
quedan fijos en él, de forma que estos tres factores más el factor IV que es el
calcio forman el complejo pro-trombinasa que convertirá la protrombina en
trombina.
79 por lo antes dicho
podemos asegurar que la coagulación se da solamente sobre la plaqueta activada
es decir en la herida no en otra parte.
80 un detalle importante
es saber que el factor X posee fuerte carga negativa y el factor plaquetario
tres también posee fuertes carga negativa por lo que es evidente que se van a
repeler para solucionar este problema entra en juego el factor IV que es el
calcio, un ión positivo. El calcio como es evidente se encuentra en el torrente
sanguíneo. Algunas sustancias como el citrato son anticogulantes porque funcionan como quelantes del calcio de forma que el complejo pro-trombinasa no se forma.
81 cuando la protrombina
entra en contacto con el complejo pro-trombinasa se convierte en trombina al
perder el péptido residual.
82 cuando las plaquetas se
activan ellas mueren porque se reinicia un proceso de apoptosis que empeso cuando la plaqueta se desprendio del megacariocito. Uno de los
procesos que se llevan a cabo durante esta apoptosis es la exposición del
factor plaquetaroa tres. En la actulidad se sabe que la enzima principal encargada de producir estos cambios en el factor plaquetario tres, a la plaqueta es la llamada ESCRAMBLASE.


83 para que el sistema de
factores sea más eficiente estos factores tienen que tener fuertes cargas
negativas. Un caso es como ya dijimos el factor X.
84 el órgano que está
encargado de hacer que estos factores adquieran cargas negativas es el hígado. Primero fabrica los factores de coagulación, los vierte
en el torrente sanguíneo y después los vuelve a recapturar para darles esta
carga negativa necesaria para que inter-actúen los factores de coagulación
correctamente.


85 el proceso por el cual
se hace posible que los factores de coagulación tengan fuerte carga negativa se
llama carboxilacion ( agregación de moléculas de carbono) y se da como ya
dijimos en el hígado. Este proceso de carboxilación
de los factores de coagulación es dependiente de la vitamina K porque interviene directamente en la carboxilación. La
ganancia de cargas negativa por parte de los factores es dependiente de la
vitamina K.

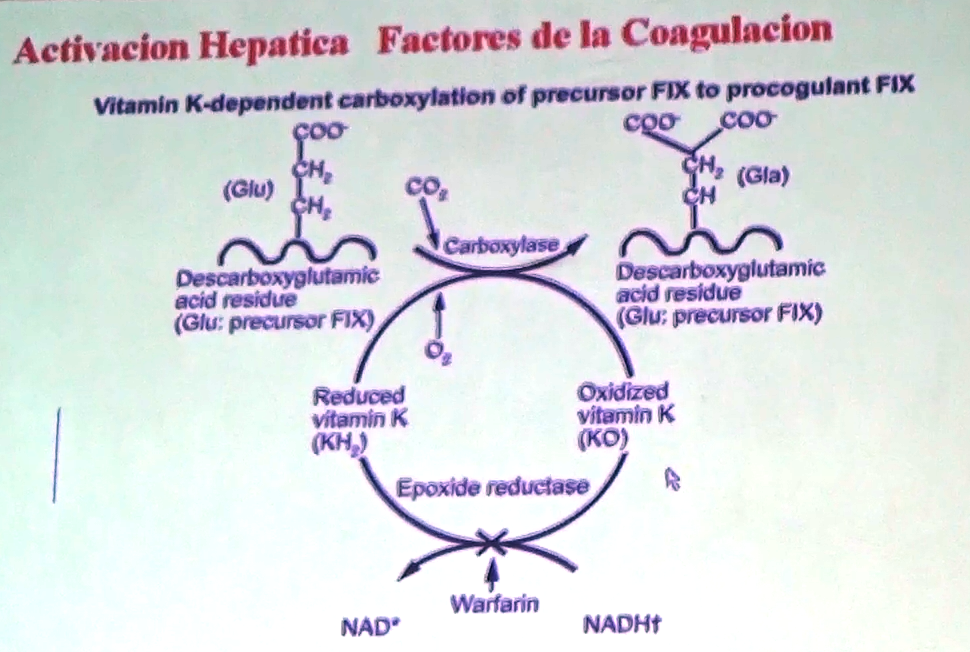
86 No todos los factores van a necesitar este proceso de carboxilación, a estos se les denomina factores
dependientes de la vitamina K uno de ellos es el X como ya sabemos.
87 el factor V es activado
por la trombina.
88 para activar al factor
X se procede a través de la vía intrínseca y de la vía extrínseca.


VÍA EXTRÍNSECA
89 la vía intrínseca se inicia con la
ruptura de las membranas de las células que sufren el trauma porque en estas
membranas se encuentra el factor tisular. Este factor no se encuentra en las
membranas de los elementos que existen en el torrente sanguíneo incluido
el endotelio. Es decir ni los glóbulos blancos ni los rojos ni las plaquetas y
tampoco el endotelio tienen en sus membranas el factor tisular, en todas las
demás membranas celulares del cuerpo si
se puede encontrar el factor tisular.


90 cuando se expone el
factor tisular este se une al FACTOR VII que siempre está activado y los dos
activan al factor X.
91 debemos observar que el
factor VII se tiene que acercar a la membrana de la misma forma en que lo hacia
el factor X a la membrana de la plaqueta. Entonces el factor VII también es dependiente de la vitamina K.
92 a la vía extrínseca se
le conoce con el nombre de la VIA FISIOLOGICA EXTERNA. Se denomina así porque
es externa al vaso sanguíneo y se produce por un lesión en los tejidos del
organismo.






1 la coagulación podía
seguir la vía intrínseca (colágeno del tejido sub-endotelial) y la vía extrínseca
(tejido lesionado) estas dos vías
coincidían en la vía común.
2 las dos vías tanto la
intrínseca como la extrínseca activan la protrombina y la convierten en
trombina. De forma que la trombina convierte o activa el fibrinógeno en
fibrina.
3 sabemos que la trombina
además de activar el fibrinógeno para que se convierta en fibrina activa
también al factor XIII, que es el factor estabilizador de la fibrina. De forma
que este factor activado hace posible que las uniones de fibrina sean
irreversibles es decir no se despolimerise convirtiéndose en fibrinógeno
nuevamente. Esto es así por las uniones de fibrina son enlaces que permiten una
reacción inversa cuando actúa el factor XIII forma enlaces covalentes
irreversibles de forma que la fibrina ya no se puede convertir en fibrinógeno
nunca más.
4 debemos reconocer y esto
es muy importante que el factor X puede ser activado tanto por la vía
intrínseca como por la vía extrínseca.
5 recordemos que el factor
X se activa por la vía extrínseca por acción del factor tisular que se
encuentra en la membrana rota de la célula lesionada más el factor siete que conjuntamente activaran al factor X que activado formará parte del
complejo protrombinasa para activar a la protrombina y convertirla en trombina.
Es por este tipo de proceso que esta vía es conocida como la vía extrínseca,
justamente debido a que es iniciada por un agente por una influencia externa.
6 la vía extrínseca es
conocida también como la vía fisiológica porque se presenta como es debido en
el momento de una lesión para no `permitir una hemorragia. Además el coagulo se
forma sobre el factor plaquetario III que se encuentra en la membrana de la
plaqueta que se encuentra en la zona de la lesión, es decir todo este proceso
ocurre en la herida y no en otra parte.
7 el factor VII también es
dependiente de la vitamina K
VÍA INTRÍNSECA
VÍA INTRÍNSECA
8 el factor X también se puede activar por la vía
intrínseca con la formación de un complejo muy similar al complejo
pro-trombinasa de la vía extrínseca. Que en vez del factor X se encuentra el
factor IX y en vez del factor V se encuentra el factor VIII. En este caso el X
sería la enzima mientras que el factor VIII es la coenzima. también se
encuentra el calcio y el factor plaquetario III.
9 sabemos que el factor IX
era activado por el factor XI y el factor XI por el factor XII, este último es
activado por una superficie activa que posee grande cargas negativas.
10 como vemos el factor IX
también se tiene que posicionar sobre el factor plaquetario III y ya sabemos
que esta mediado por el calcio, un ión positivo. Por lo visto también el factor
IX es dependiente de la vitamina K
11 en conclusión diremos
que los factores dependientes de la vitamina K son:
- X 10
- IX 9
- VII 7
- II 2
12 en la parte teórica la
coagulación no tiene dos vías pero en la parte práctica los análisis de
laboratorio cumplen de manera convincente este presupuesto.

Los análisis de laboratorio evalúan sus parámetros para el diagnóstico en base a las dos vías. TPT(tiempo parcial de tromboplastina) evalúa como veremos la vía intrínseca. El TP (tiempo de protrombina) es evaluado sobre la vía extrínseca. De acuerdo al resultado de estas pruebas se le da o no un tratamiento a los pacientes. En conclusión diremos que la teoría de las dos vías de la coagulación son útiles en los análisis clínicos.


Los análisis de laboratorio evalúan sus parámetros para el diagnóstico en base a las dos vías. TPT(tiempo parcial de tromboplastina) evalúa como veremos la vía intrínseca. El TP (tiempo de protrombina) es evaluado sobre la vía extrínseca. De acuerdo al resultado de estas pruebas se le da o no un tratamiento a los pacientes. En conclusión diremos que la teoría de las dos vías de la coagulación son útiles en los análisis clínicos.

13 uno de los problemas en
la teoría de los factores de la coagulación es que el factor XII o factor de
Hageman no lo tienen algunas personas y sin embargo no tienen problemas de
coagulación secundaria o hemostasia secundaria. Se piensa también que el factor
tisular junto al factor VII son los que pueden activar al factor IX.
FASE DE FIBRINOLISIS
hemostasia terciaria
FASE DE FIBRINOLISIS
hemostasia terciaria
14 una vez que el tejido
endotelial ha sido reparado el coagulo de fibrina es un estorbo para el tejido
por lo cual tiene que ser retirado.
15 el proceso por el cual
el coagulo de fibrina es eliminado se lleva a cabo por enzimas especializadas
de forma que este coagulo pueda ser desensamblado y luego fagocitado por los
macrófagos.
16 el PLASTINOGENO, que es
otro factor se adhiere a la fibrina apenas esta se forma.


17 los tejidos lesionados
y los tejidos en reparación activan al activador tisular del plastinogeno que
se va a unir al plastinogeno activándolo y convirtiéndolo en PLASTINA que es la
enzima con la capacidad para disolver a la fibrina. A este proceso le denominamos fibrinólisis.


18 el proceso de
fibrinolisis es muy eficiente porque el
platinogeno ya está junto a la fibrina de forma que el activador tisular de
plastinogeno lo encuentra fácilmente.
19 los productos de
degradación de la fibrina (PDF)
20 en el laboratorio se
puede medir la concentración de PDF de manera que se puede determinar, si en
caso estuviera elevada esta concentración la formación de trombos.
21 como ya dijimos todos
los procesos de la coagulación se presentan al mismo tiempo. La diferencia está
en que al principio de la coagulación se intensifica la formación de fibrina
mientras que al final se intensifica la fibrinólisis.

LA SANGRE
La sangre es un fluido acuoso con elementos sólidos en suspensión. presenta un movimiento pulsatil y perpetuo. se encuentra contenido dentro de los vasos sanguíneos.

22 la función más importante de la sangre es el transporte de oxigeno y de CO2 cualquier otra función puede fallar pero solo tendrá un efecto a largo plazo. La falta de oxígeno es sin embargo letal para el organismos.


22 la función más importante de la sangre es el transporte de oxigeno y de CO2 cualquier otra función puede fallar pero solo tendrá un efecto a largo plazo. La falta de oxígeno es sin embargo letal para el organismos.

23 la sangre en el tubo de
ensayo se puede estudiar centrifugándola por un tiempo y determinado así la
formación de la parte de los elementos formes(fase) que es una sustancia roja y
el plasma que es una sustancia amarilla(fase). los elementos formes constituyen
el 45 % del volumen total mientras que el plasma constituye el 55%. También
podemos llegar a observar una inter-fase entre la fase de glóbulos rojos y la
fase del plasma que es la fase leucocitaria compuesta por leucocitos y
plaquetas.




24 cuando la sangre se
extrae con un anticoagulante se puede observar efectivamente el plasma mientras
que si no se extrae con anti-coagulante se forma el suero.
25 la diferencia entre el
suero y el plasma es la presencia de los factores de coagulación. Si se extrae
una muestra de sangre en un tubo con anticoagulante lo que tendremos es plasma
mientras que si se recolecta la sangre en un tubo sin anticoagulante la muestra
después de un tiempo presentará el suero. Un tubo
con anticoagulante permitirá que la fase del plasma contenga los factores de
coagulación ya que se encuentran inactivos debido al anticoagulante mientras
que en el tubo sin anticoagulante los factores de coagulación ya no se
encuentran presentes porque estos han reaccionado formando complejos que ya no
se encuentran en el suero debido a su densidad por ello el plasma se queda sin
estos factores y a este plasma sin factores le denominamos suero.
PLASMA
PLASMA
26 el 90 % del plasma es
agua mientras que el 9% esta formado por diferentes proteínas el restante 1 %
son los otros elementos del suero como iones, carbohidratos o lípidos.
27 la albumina y la
globulina son las dos proteínas que se encuentran en más abundancia en el
suero.


28 la concentración de
proteínas tiene un valor que va de 3-8 mg/% y siempre la concentración de
albumina es mayor que la de globulinas.
29 las globulinas se
dividen en
-alfa 1
-alfa 2
-beta
-gamma


30 las concentraciones de
los diferentes tipos de proteínas se determinan en los laboratorios por
electroforesis, dividiendo a las proteínas a través de su carga eléctrica.
Recordemos también que la proporción o el cociente de la albumina con las globulinas es de
1.5 a 2.7. por ello la albúmina siempre será mayor que las globulinas.
 Observamos en la imagen de arriba que la proporción de albumina es diferente dependiendo de la edad del individuo.
Observamos en la imagen de arriba que la proporción de albumina es diferente dependiendo de la edad del individuo.

ELEMENTOS FORMES Y CELULARES
31 hablamos de la fase de los elementos formes porque la mayor parte de esta fase está formada por glóbulos rojos y estas no son consideradas células porque no tienen núcleo y tampoco organelas.

32 los elementos celulares
de la sangre son una fracción denominada leucocitos formados por células
encargadas exclusivamente de la defensa
del organismo contra agentes patógenos, es decir son los encargados de
organizar la respuesta inmune del organismo.


33 dentro de el grupo
células de la sangre encontramos a los polimorfonucleres que se les denomina de
esa forma porque poseen un núcleo con varios lóbulo.
34 el otro grupo serían
los mono-mornucleres, es decir los que tienen u
núcleo si lóbulos tales son
los monocitos y los linfocitos.
35 dentro del grupo de los
polimorfonucleres encontramos a los basófilos, presentan una tendencia a la
coloración azul, los eocinofilos que presentan una tendencia a la coloración
rojiza y los neutrófilos que no presentan una tendencia muy marcada por los dos
colores.
36 los monocitos se
encuentran en la sangre pero cuando estos migran hacia los tejidos e denominan
macrófagos.
37 como dijimos en la
inter-fase entre la fase de los elementos formes y la fase del plasma
encontramos a la fase leucocitaria donde encontramos como su nombre lo dice a
los leucocitos y por sobre estos leucocitos podemos hallar a las plaquetas.


38 esta disposición de
arriba hacia abajo: plasma-plaquetas-leucocitos-eritrocitos se da por la
densidad de estos elementos.
39 por cada milímetro
cubico encontramos de 4-5 millones de eritrocitos mientras que solo encontramos
de 5-10 mil leucocitos.


40 el volumen de los
elementos formes sobre el volumen total
es el hematocrito termina siendo una expresión de la cantidad de glóbulos rojos
que se encuentran en la sangre. por lo tanto a mayor hematocrito mayor cantidad
de glóbulos rojos.
41 los glóbulos blancos
solo representan el 0.4% del volumen total.
HEMATOPOYESIS
HEMATOPOYESIS
42 la formación de la
sangre se da por un proceso que se denomina hematopoyesis.
43 todas los elementos que
conforman la fase de elementos formes y la fase leucocitaria provienen, por el proceso de hematopoyesis de
una célula madre denominada STEM CELL o célula troncal.
44 la hematopoyesis es un
proceso muy activo y complejo además requiere de factores de crecimiento como: eritropoyetina, trompoyetina, citoquinas, quimioquinas, etc. Se realiza en la médula ósea roja.
45 todas las células de la
sangre provienen de una célula madre también denominada célula multipotecial


46 la stem cell gracias al proceso de diferenciación y transformación se convierte primero en una
célula mieloide-linfoide a la vez y luego en su proceso de diferenciación
solamente en mieloide y por otro lado en linfoide. Las células linfoides dan
origen a los linfocitos mientras que las células mieloides dan origen a las
demás células.Ni las células mieloides puras ni las linfoides puras pueden dar marcha atrás en su proceso de diferenciación.
47 las células madre tienen la capacidad de autorregeneración. Esto quiere decir que antes de
dividirse y diferenciarse en un tipo especifico de células generan una
célula idéntica a ella para que en lo sucesivo haga lo mismo. Por ello podemos
decir que las células madres son células progenitoras con capacidad de
auto-regeneración o renovación.


48 las células madres son
células progenitoras pero no todas las células progenitoras son células madres.
Recordemos que una célula progenitora es aquella que puede dar progenie pero
eso no quiere decir que pueda renovarse.
49 a medida que una célula
se va dividiendo y diferenciando va perdiendo su capacidad de regeneración. Las células con menos divisiones, es decir, las que constituyen una reserva de células madres son las más indiferenciadas y las más
primitivas.
50 un detalle importante
de recordar es que las células resultantes de la mitosis son idénticas mientras
que las células resultantes de una meiosis son diferentes.






51 en el organismo tenemos
en principio las células madres TOTIPOTENTES estas células tienen la capacidad
de generar un organismo completo. Es decir generan al individuo propiamente
dicho incluido todos los tejidos extraembrionarios como la placenta. Estas células
son las que se pueden clonar. Podemos encontrar células madre totipotentes
hasta la fase embrionaria de mórula.
52 una vez que se ha pasado la
fase de mórula las células pierden su capacidad de totipotencialidad y
solamente serán capaces de generar tejidos embrionarios o tejidos
extraembrionarios pero no los dos la vez
a esta célula madre se le conoce como PLURIPOTENTE.


53 las células pluripotentes dan origen en su curso normal de diferenciación al mesodermo, ectodermo y
endodermo. Por lo que las células de estos linajes solamente serán capaces de
formar tejidos que tengan origen mesodérmico endodérmico o ectodérmico. A este
tipo de células madres se les denomina células MULTIPOTENTES.


54 como vemos las células
madres van perdiendo potencialidad a medida que se van dividiendo y
diferenciándose.
55 observemos que las
células multipotentes son capaces de generar solamente un linaje en especifico. Solamente podrán generar tejidos que tienen un origen común
mesodérmico, ectodérmico y endodérmico.
56 La
célula madre hematopoyética es una célula multipotente que solamente puede dar
origen a un linaje especifico de tejidos. En este caso es la célula que da
origen a los tejidos derivados del mesodermo.


Consideremos que son las células madres multipotententes las que se usan para las terapias celulares.
57 las células hematopoyéticas o multipotentes de la sangre pueden generar las células sanguíneas y al hacerlo pierden su potencialidad pero ganan especificidad o diferenciación. De esta forma ganan funcionalidad dentro del organismo. Por ello en un primer proceso las células madres se convierten en células progenitoras es decir que ya no tienen capacidad de auto-renovarse.
57 las células hematopoyéticas o multipotentes de la sangre pueden generar las células sanguíneas y al hacerlo pierden su potencialidad pero ganan especificidad o diferenciación. De esta forma ganan funcionalidad dentro del organismo. Por ello en un primer proceso las células madres se convierten en células progenitoras es decir que ya no tienen capacidad de auto-renovarse.
58 las células
progenitoras como bien sabemos tienen la capacidad de dividirse es decir de
generar una progenie pero ya no de auto-renovarse.
59 En seguida las células
progenitoras se dividen generando una progenie de células precursoras las
cuales han perdido la capacidad de dividirse pero son mucho más especializadas
o están en una vía en la que adquieren capacidades muy especificas dentro del
organismo.
60 las células
diferenciadas son células plenamente funcionales para el organismo las cuales
han perdido totalmente la capacidad de auto-renovarse y de dividirse.



61 en este proceso de la hematopoyesis podemos reconocer así la naturaleza de cada célula o etapa de la hematopoyesis:
-célula multipotencial o
hematopoyética: STEM CELL hematopoyetica
-célula progenitora:
unidad formadora de colonias linfoide y mieloide. Y también las células
formadoras de colonias linfoides y las células de la unidad formadora de
colonias mieloides.
-célula precursora: son los blastos. Como por ejemplo el
eritroblasto o el linfoblasto o megacarioblasto.
-célula diferenciada: son
las que tienen funcionalidad completa tales como el eritrocito, el linfocito o
el macrófago.
62 las células
multi-potenciales para conservar su naturaleza entran en el G0 una etapa de
quiescencia para la célula. Cuando se presentan los factores estimulantes
necesarios la célula madre sale de la etapa G0 para entrar de nuevo en
actividad y dividirse con el propósito de generar células especializadas en la
medida en que el organismo lo requiera. A parte de las células madres existen
otras células que también se encuentran en estado quiescente, ellas son por
ejemplo la neurona o el hepatocito que si se presentan las circunstancias
adecuadas pueden dividirse también.


63 las STEM CELL son
células plenamente indiferenciadas y por ese motivo no tienen marcadores de
diferenciación como lo tienen las células de los tejidos normalmente que son
células diferenciadas.
64 los marcadores de
diferenciación son moléculas que definen a un tejido. Por ejemplo el músculo
esquelético tiene una molécula denominada miogenina, esta molécula no la
presenta ninguna otra célula del cuerpo más que las células del musculo
esquelético.


65 la célula madre
totipotencial es la célula madre que no tienen ningún marcador de manera que
todavía no ha elegido ningún camino pero cuando esta se divide toda la decisión
de seguir un camino especifico convirtiéndose así en una célula madre
multipotencial. En nuestro caso sería la célula madre hematopoyética que ya
tiene un primer marcador de diferenciación denominado CD-34 por lo cual este
tipo de célula multipotencial solamente puede generar células sanguíneas.




66 carece del complejo
mayor de histocompatibilidad tipo 1(MHC-1) que lo hace invisible al sistema
inmunológico de manera que no puede ser atacado por las defensas del organismo.
FUNCIONES DE LOS FACTORES
ESTIMULANTES (CITOQUINAS) HEMATOPOYETICAS
67 sirven para que la
célula madre pueda seguir uno de cinco caminos:
-supervivencia: inhibición de la apoptosis. Si la célula no tiene el factor estimulante entra en apoptosis.
-proliferación: inducción
mitosis
-diferenciación celular
-inducción de la
maduración
-estimulación
funcional


68 la interleuquina 5 da origen a los eocinofilos por ello es considerada un factor
estimulante de la hematopoyesis muy especifico, mientras que la IL3 se encuentra como factor estimulante de muchos elementos de la sangre.


69 los factores
estimulantes se pueden dividir en tres grupos:
-Iniciación: actúa a
nivel de las STEM CELL (IL3,IL11,IL12).
-acción intermedia: actúan
a nivel de las células progenitoras(IL3,IL4).
-acción tardía: actúan a
nivel de las células precursoras(trombopoyetina y eritropoyetina).
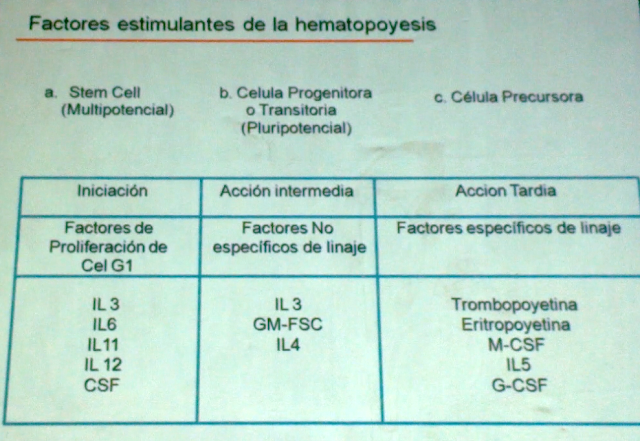

70 casi todos los órganos
pueden liberar factores estimulantes entre ellos el riñón, el hígado y los
pulmones.
71 las células sanguíneas
se producen en la médula ósea roja.


72 a los 5 o 6 años la
médula ósea roja se convierte en médula ósea amarilla es decir se reemplaza por grasa.
73 los focos
hematopoyéticos están en casi todos los huesos pero sobre todo en las vértebras
y en la pelvis. En ellos se realiza más del 50% de la producción de las células
sanguíneas.




74 en la médula ósea roja
encontraremos:
-75% está formado por
leucocitos
-25% está formado por
eritrocitos
75 estos porcentaje casi
son los opuestos a los que se encuentran en la sangre. Se debe a que los
eritrocitos viven 120 días mientras que los leucocitos viven solamente algunas
horas de forma que la producción de leucocitos debe ser más activa a nivel de
la médula.


76 en la médula hay una
célula madre por cada 10000 células. Y de vez en cuando estas células madres
pueden llegar a salir al torrente sanguíneo pero no es un proceso normal y es
poco corriente.
77 el nicho adecuado es el
ambiente en el cual la célula madre hematopoyética encuentra un ambiente
adecuado para su división y especialización debido a que esta en constante
interacción con los factores estimulantes.


78 existen dos tipos de
factores estimulantes a este nivel:
-los factores derivados
del estroma (tejido conectivo en la médula) esta alberga a cinco tipos
celulares que son imprescindibles para que se realice el proceso de la
hematopoyesis:
-macrofagos
-fibroblastos que forman
la matriz conectiva
-los osteoclastos
-osteoblastos
-osteoblastos
-adipocitos
Sabemos que si falta
alguno de estos cinco tipos celulares no se da la hematopoyeis.

-los factores derivados
del osteoblasto
79 por lo visto la
hematopoyesis es extravascular


Obajo observamos como se extrae parte de la médula ósea roja de la cresta iliaca para su estudio clínico:

80 Volver a repoblar la médula ósea, después de una irradiación para combatir el cáncer, se requiere un trasplante de médula ósea. Este trasplante no es una operación. Se hace mediante el filtrado de la sangre del donante tratando de capturar las células madres que se han desprendido hacia el torrente sanguíneo. las células madre del donante son transfundidas en la médula ósea del paciente. De esta manera se facilita la repoblación de las células madre reiniciándose la hematopoyesis.


80 Volver a repoblar la médula ósea, después de una irradiación para combatir el cáncer, se requiere un trasplante de médula ósea. Este trasplante no es una operación. Se hace mediante el filtrado de la sangre del donante tratando de capturar las células madres que se han desprendido hacia el torrente sanguíneo. las células madre del donante son transfundidas en la médula ósea del paciente. De esta manera se facilita la repoblación de las células madre reiniciándose la hematopoyesis.

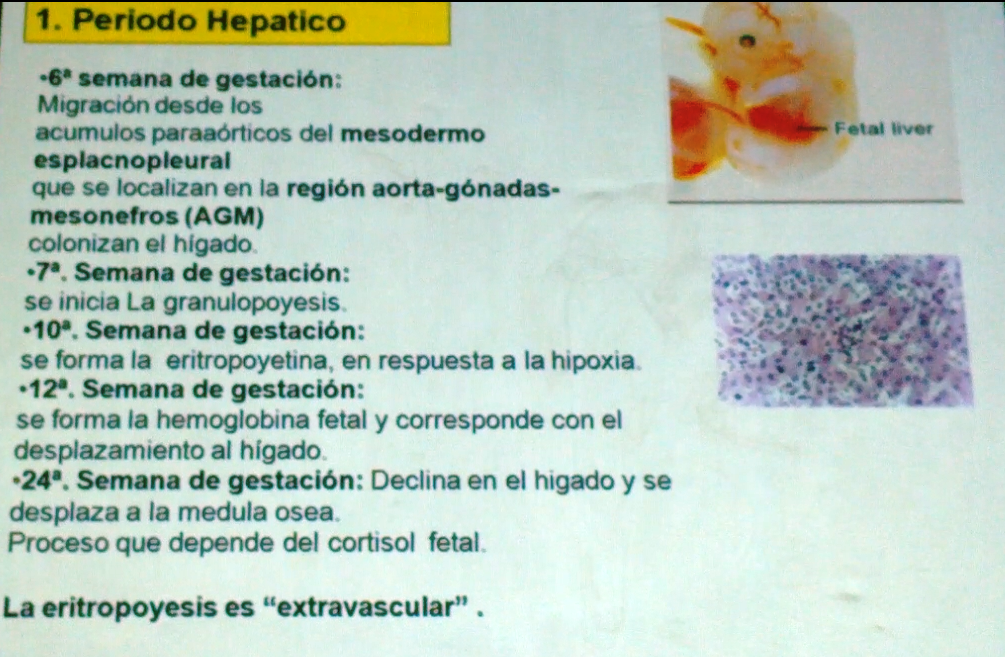
81 en principio durante el periodo mesoblastico la
formación de la sangre se da en el saco vitelino,es decir en el embrión, donde solo se forman eritrocitos y todavía no se realiza la formación de glóbulos blancos. Posteriormente la formación de la sangre pasa a ser función del hígado, en el feto, por lo que se denomina el periodo hepático. En ciertos procesos patológicos en el adulto, cuando no se puede realizar la hematopoyesis en la médula ósea roja, esta función es retomada por el hígado en un proceso que denominamos metaplasia mieloide. Esto solamente se puede presentar porque ya antes el hígado realizó esta función. Y finalmente después de la formación de los huesos, en la médula ósea roja.





83 la médula ósea roja incrementa su producción alrededor de los dos años. luego alrededor de los 5 o 6 años empieza a menguar la producción. la disminución de la producción de células sanguíneas se presenta porque la médula ósea roja es reemplazada por médula ósea amarilla, es decir grasa. La declinación de la producción se presenta de forma más notable en los huesos largos en donde se produce un reemplazo por grasa. Los huesos axiales mantienen su actividad a lo largo del tiempo y son precisamente los lugares en donde se realizará la hematopoyesis a lo largo de toda la vida.


En la imagen de arriba observamos una gráfica edad versus producción de células sanguíneas. Las curvas representan la actividad de los centros de producción(médula ósea roja) de células sanguíneas. Los primeros dos meses la producción de eritrocitos se presenta en el saco vitelino, después esta función es responsabilidad del hígado y también del bazo. Alrededor del cuarto mes se da la producción de sangre en la médula ósea roja. finalmente alrededor de los 5-6 años se inicia el reemplazo de la médula ósea roja por amarilla declinando la producción de forma notable en los huesos largos.
ERITROPOYESIS
85 la eritropoyesis es la
formación de los glóbulos rojos.
86 el proceso de la
eritropoyesis comienza cuando la STEM CELL se divide y genera la primera
célula progenitora que es la CFU-ML que genera la segunda célula progenitora
que es la CFU-M que genera la unidad formadora de grandes colonias de
eritoblastos (BFU-E) que aún es una célula progenitora con capacidad para
regenerarse.
88 la célula BFU-E genera
a la célula precursora que es el eritoblasto
89 El eritroblasto genera finalmente a la célula diferenciada
que llamamos eritrocito.


90 eritropoyesis:

-Células blástica:
STEM CELL
-Células progenitoras:
recordemos que a este nivel se presenta la influencia de la eritropoyetina:
*Precursor granulo
eritroide macrofagico y megacariocitico. esta es la unidad formadora de
colonias mieloide
*BFU-E: unidad formadora de grandes
colonias. se sabe que entre 10 Y EL 20 % están en el ciclo celular. Forman entre 30 mil y 40 mil celulas al día. depende fuertemente de las citoquinas. Aun pueden ser renovados.
*CFU-E: unidad formadora de colonias
(pequeñas).Son progenitoras con una gran diferenciación. del 60-80% entran al ciclo. Forman de 8-65 celulas en 7 días. No se encuentra en sangre periférica y no pueden ser renovados. depende fuertemente de la erotropoyetina.
-Células precursoras:


*Proeritroblasto: mide de 20-25
micras
*Eritroblasto basófilo: en esta etapa se da una gran producción de proteínas(para la producción de hemoglobina) y por lo tanto existen muchos ribosomas y ácido nucleico por lo cual son basófilos (color azulado) miden de 15 a 20 micras.
*Eritroblasto basófilo: en esta etapa se da una gran producción de proteínas(para la producción de hemoglobina) y por lo tanto existen muchos ribosomas y ácido nucleico por lo cual son basófilos (color azulado) miden de 15 a 20 micras.
*Eritroblasto
policromatofilo: en esta etapa ya se esta produciendo la hemoglobina que es
rojisa por lo tanto esta celula presentara una variedad(entre rojiso y azulado) de colores al
microscopio. Mide de 8-12 micras
*Eritoblasto ortocromatico (verdadero color):también llamados normoblastos. En esta etapa ya se encuentra sintetizada la mayor parte de hemoglobina por lo
tanto asume su verdadero color rojizo. Mide de 7-10 micras.
91 observamos que en el
proceso de maduración de la célula precursora el núcleo se va empaquetando o
condensando de forma que queda muy reducido y al final de su etapa de célula
precursora, cuando ya ha sintetizado la hemoglobina necesaria para su función, el núcleo es expulsado del citoplasma.
92 el reticulocito es una
célula de 8 micras a la que le falta muy poco para convertirse en eritrocito plenamente funcional
solo le queda por expulsar fragmentos de ácido nucleico que están en su
citoplasma los cuales le hacen ver al microscopio como una red dentro de esta
célula.
93 finalmente el
reticulocito se convierte en glóbulo rojo adoptando su forma normal de disco
bicóncavo de 7 micras.
94 por todo lo antes dicho
el eritrocito no es una célula sino un derivado de una célula que era el
eritroblasto. Por ello solo lo consideramos un elemento forme.


95 en un hemograma es
normal encontrar reticulocitos en un porcentaje de 0.5 -1.5 % mientras que los
glóbulos rojos están en el porcentaje de 98 %.
96 el reticulocito es un indicador de la producción de glóbulos rojos en la médula ósea. Pueden presentarse dos casos distintos:
96 el reticulocito es un indicador de la producción de glóbulos rojos en la médula ósea. Pueden presentarse dos casos distintos:
-que se presente anemia
con reticulocitos altos: esto indicaría que la médula ósea esta produciendo
mucha más cantidad de elementos en su afán de compensar la falta de eritrocitos
por lo cual deducimos que se están perdiendo glóbulos rojos por una posible
hemorragia.
-que se presente anemia
con reticulocitos bajos: quiere decir que no se están produciendo glóbulos
rojos por lo que se presenta la anemia
97 si se llegan a
encontrar eritroblastos ortocromaticos podemos saber que la médula está haciendo
un esfuerzo extremo por producir más células sanguíneas de manera que libera a
estos elementos aun cuando presentan núcleos.
98 el reticulocito tiene
que expulsar liquido de su citoplasma para convertirse en glóbulo rojo
plenamente funcional. Esto quiere decir que la forma típica de disco bicóncavo
del eritrocito es como una bolsa a medio llenar. Esta condición le permite al
eritrocito poder adaptarse al diámetro reducido de los capilares. Esto indica
que es importante que se termine la maduración del eritrocito.
Recordemos que un globulo rojo vive 120 días:

Recordemos que un globulo rojo vive 120 días:

99 la función principal de
la eritropoyetina es inhibir la apoptosis de la célula madre de forma que el
resultado es el incremento de la producción de eritrocitos.


100 la eritropoyetina se
produce en las células intersticiales del riñón porque es ahí donde se
encuentran los sensores de oxígeno. Se incrementa la concentración de
eritropoyetina cuando se detecta que existen niveles bajos de oxigeno en la
sangre arterial.


101 podemos decir que la
eritropoyetina es sensible a los niveles de oxigeno y no a la cantidad de
glóbulos rojos.
102 los sensores de
oxigeno se encuentran en el riñón porque es el órgano más activo del cuerpo por lo que consume gran cantidad de oxigeno en comparación con otros órganos. Si hay deficiencia de oxigeno en
los riñones entonces es evidente que se necesita aumentar los niveles de
oxigeno en sangre. Si se da una condición de hipoxia el
primer tejido que sufre estrés es el tejido renal.
103 la eritropoyetina
también tiene una función angiogenetica y vasculogenetica además también juega
un papel en el tejido nervioso como neuroprotector evitando la apoptosis.

104 en caso de una insuficiencia renal probablemente sufra de anemia severa

104 en caso de una insuficiencia renal probablemente sufra de anemia severa
105 la eritropoyetina
actúa por el factor inducido por hipoxia.


106 cuando las
concentraciones de oxigeno en sangre son adecuadas el factor inducido por
hipoxia se degrada por acción de la ubiquitina de manera que la producción de
eritropoyetina es normal. En caso contrario el factor indicido por hipoxia
acciona la producción superior de eritropoyetina
107 también existen
factores inhibidores de la eritropoyetina como el factor de necrosis tumoral y otras moléculas que son productos de la inflamación. Muchas enfermedades inflamatorias como la artritis pueden producir anemia.


HEMOGRAMA

108 dentro de la variables que se hacen en un hemograma tenemos:
-volumen corpuscular
medio: Es el tamaño promedio de un glóbulo rojo que se encuentra entre 80-96 fL.

Concentración media de hemoglobina:es la concentración de hemoglobina en cada glóbulo rojo.
109 si la hemoglobina(molécula soluble) estuviera disuelta en el plasma se perdería por los riñones debido a su peso molecular. Por esto no puede estar libre.

109 si la hemoglobina(molécula soluble) estuviera disuelta en el plasma se perdería por los riñones debido a su peso molecular. Por esto no puede estar libre.
110 la hemoglobina fuera
del eritrocito es altamente toxica. Por eso cuando se da la lisis masiva de
glóbulos rojos la hemoglobina se libera y se difunde a través del endotelio a
nivel del glomérulo renal de forma que ingresara en los tubulos renales causando
la necrosis del epitelio del túbulo.
111 un accidente cerebro
vascular hemorrágico es mucho más dañino para las neuronas que uno isquémico. Esto se da debido a que la hemoglobina entra en contacto con las
neuronas causando la muerte de estas.
112 El glóbulo rojo cuenta con un sistema enzimático limitado para evitar
la oxidación de la membrana del eritrocito por los radicales libres. Cuando este mecanismo deja de
funcionar la membrana del eritrocito pierde su característica más importante
que es su flexibilidad de manera que al pasar por un capilar estrecho ya no puede adaptarse a la dimensión del capilar. Esto ocasiona la
destrucción del eritrocito. El eritrocito tiene una vida limitada
porque no tiene la capacidad de sintetizar y regenerar su propia membrana por
mucho tiempo.


113 cuando el eritrocito
pasa por el bazo se encuentra con capilares muy finos de forma que este se
queda atascado y es detectado y fagocitado por los macrófagos del bazo.
INMUNIDAD
1 Es la acción mediante la cual el organismo se defiende de los elementos patógenos.
2 la stem cell se diferencia en dos grande líneas:
-línea mieloide:
constituido por los monocitos y los polimorfonucleres(neutrófilos, eocinofilos
y basófilos) . También están incluidos en este grupo las plaquetas y los
eritrocitos.
-línea linfoide: estas
células van a migrar para su proceso de maduración. A este grupo pertenecen los
linfocitos.


3 los polimorfonucleares
solamente se distinguen por la coloración de sus gránulos al microscopio. Pero
el neutrófilo está más emparentado con el monocito que con los otros dos
polimorfonucleares.
4 el neutrófilo y el
monocito están muy emparentados de forma que también en su función son bastante
parecidos. Los dos son fagocitos y cumplen esta función en el organismo.
5 recordemos que todas las
células del cuerpo pueden fagocitar pero los monocitos y los neutrófilos están
diseñados para eso. Es decir son fagocitos profesionales.
6 los leucocitos se
diferencian en cinco líneas:
-Neutrófilos
Abastonados: 0-4 %
Segmentados : 50-70 %
-Eosinofilos : 2-4 %
-Basófilos: 0-1%
-Monocitos : 2-8 %
-Linfocitos : 25-40 %


7 por lo visto anteriormente los neutrófilos son
los de mayor concentración en la sangre.
LEUCOPOYESIS
8 es la formación de los
glóbulos blancos
9 STEM CELL que se
transforma en FCU-LM(unidad formadora de colonias linfoide y mieloide) y estas
a su vez producen dos ramas la CFU-M(unidad formadora de colonias mieloide) y la
CFU-L(unidad formadora de colonias linfoide)

10 el proceso es como sigue:
stem cell generan células progenitoras y estas últimas a las células precursoras y finalmente a células diferenciadas que ya no se dividen
11 la diferenciación de los leucocitos depende
de una serie de factores estimulantes y estos se denominas citoquinas.

Las citoquinas tienen cuatro propiedades :

Las citoquinas tienen cuatro propiedades :
-pleiotropismo: como
ejemplo tomaremos a la interleuquina4 (IL4) que ejerce influencia sobre varios
tipos de células y estos realizan diferentes funciones a su vez. A esta
propiedad de generar acciones diferentes se le denomina pleiotropismo.
-redundancia: diferentes
interleuquinas tienen un mismo efecto sobre una célula por ejemplo la
proliferación del linfocito B
-sinergia: quiere decir
que solo se dará un efecto en las células con la participación activa de dos
citoquinas.
- antagonismo: dos
citoquinas producen efectos contrarios sobre las células


12 la inmunidad nos sirve
para defendernos de los diferentes agentes extraños que pueden ingresar al
organismo.
13 la inmunidad empieza
por el reconocimiento de los agentes que entran en el organismo de forma que
constantemente hay un reconocimiento de lo que es propio y lo extraño. Todo aquello que sea reconocido como propio será considerado inocuo y lo extraño como nocivo.


14 este reconocimiento es
molecular. El leucocito posee en su membrana celular receptores químicos y de
manera complementaria los agentes externos que puedan ingresar poseen
estructuras en sus membranas denominadas ligandos. De forma que se puede dar el
reconocimiento molecular.
15 si el leucocito
encuentra al ligando lo catalogará como extraño y potencialmente nocivo por lo
cual se iniciara un proceso de destrucción de este elemento y si no encuentra
este ligando lo dejara libre. Lo que hace en términos más prácticos el
leucocito, es ir palpando a los elementos que se encuentran en su recorrido y
así identificar a todos ellos.
16 en resumen la inmunidad
se basa en receptores y ligandos
17 la inmunidad la podemos
dividir en dos grandes grupos según como se formen los receptores:
-Inmunidad innata: los receptores se forman desde el nacimiento
de la célula defensiva. Esta a su vez se divide en:
*Barreras naturales
*Fagocitos complemento
-Inmunidad adaptativa: los
receptores se forman cuando la célula es ya adulta porque tiene la capacidad de
sintetizar receptores nuevos. Esta se divide a sus vez en:
*Celular
*Humoral


18 en la historia de la
evolución primero surge la inmunidad innata y después la inmunidad adaptativa.
19 la inmunidad innata
sirve para defendernos de las bacterias y por ello todas las células nucleadas
tienen inmunidad innata. Es decir poseen receptores para detectar baterías.
Recordemos que hasta las amebas tienen esta inmunidad. Esta inmunidad es muy
antigua porque las bacterias son muy antiguas también. Se piensa que los
primeros seres eucarióticos tuvieron que desarrollar este tipo de defensa
contra las bacterias. Desde tiempos remotos en la historia de la vida los seres eucarioticos aprendieron a defenderse de las bacterias nocivas. las células eucarioticas desarrollaron con el tiempo los receptores que les servían para identificar a las bacterias nocivas. Los receptores fueron transmitidos geneticamente a los descendientes de los primeros eucariotas. a lo largo de la historia de la vida los seres eucarioticos han logrado desarrollar y heredar geneticamente un repertorio amplio de receptores que garanticen una buena inmunidad innata.
20 vemos que en el ambiente no solo existen bacterias como elementos patógenos sino que surgen otros tipos de patógenos como los virus que son organismos con una alta tasa de mutaciones es decir que surgen geneticamente diferentes tipos de patógenos a un ritmo incrementado. Frente a este problema la inmunidad del organismo tiene que desarrollar nuevos receptores para destruirlos en una posterior etapa. A la inmunidad por la cual el organismo genera receptores nuevos cuando este ya es adulto se le denomina inmunidad adquirida.

20 vemos que en el ambiente no solo existen bacterias como elementos patógenos sino que surgen otros tipos de patógenos como los virus que son organismos con una alta tasa de mutaciones es decir que surgen geneticamente diferentes tipos de patógenos a un ritmo incrementado. Frente a este problema la inmunidad del organismo tiene que desarrollar nuevos receptores para destruirlos en una posterior etapa. A la inmunidad por la cual el organismo genera receptores nuevos cuando este ya es adulto se le denomina inmunidad adquirida.

21 el organismo intenta
elaborar nuevos receptores de membrana para los patógenos que son también
nuevos para el organismo pero esto no quiere decir que siempre lo haga con
éxito. Un ejemplo de ello es la gripe o influenza que mato muchas personas. Si
las personas sobreviven a esta infección entonces pasaran esos receptores a sus
descendientes por lo cual se encuentran ya inmunizados contra ese elemento de
manera que ya forma parte de su inmunidad innata.


COMPONENTES DE LA
INMUNIDAD INNATA
barreras:
-epitelios: prevenir el
ingreso de los microorganismos
-defensinas: eliminación
de microorganismos
-linfocitos
intraepiteliales: eliminación de microorganismos
células efectoras: actuan para eliminar los gérmenes.
-Neutrófilos: fagocitosis
temprana
-Macrófagos: fagocitosis,
activación e inflamación
-Células NK: eliminación
de células infectadas
proteínas efectoras: ayudan a eliminar los gérmenes
-complemento: eliminación,
opsonización
-proteína C reactiva :
opsonización
-lectina de unión a manosa
: opsonización


25 la gran diferencia
entre la inmunidad innata y la adquirida es que en la innata apenas el patógeno
entra este es reconocido y combatido mientras que en la adquirida una vez que
el patógeno entra este debe ser analizado o estudiado molecularmente, debe hacerse una plantilla de él y
seguir un proceso para generar receptores. Todo esto demora entre días y
semanas. Por ello decimos que la inmunidad innata es automática y rápida
mientras que la adquirida demora mucho más.
26 se puede decir también
que la inmunidad innata es inmediata mientras que la adaptativa tiene un
periodo de latencia.
27 dentro de la inmunidad
innata encontramos a los fagocitos neutrófilos y macrófagos mientras que en la
inmunidad adaptativa encontramos a los linfocitos B que producen los
anticuerpos y a los linfocitos T que generan los receptores de membrana.


28 la piel es una barrera
física para las patógenos
29 también se puede dar la
eliminación de microbios por sustancias antibióticas se encuentran en los
epitelios.
30 eliminación de células
infectadas y microbios por los linfocitos intra-epiteliales.


LOS RECEPTORES DE LA INMUNIDAD
31 en cuanto a LOS RECEPTORES DE LA INMUNIDAD podemos decir:
-inmunidad innata: los
receptores que presentan los fagocitos son aquellos que pueden reconocer una molécula
común en una familia de patógenos. Por ello el receptor se dice que no es
específico para un solo germen de manera que funciona de forma genérica para
todo un grupo. Por ello se dice que el fagocito busca patrones en los elementos
sospechosos.
33 los receptores de membrana de la inmunidad innata reconocen los patrones moleculares de familias de gérmenes y estos por lo general son carbohidratos. Por ejemplo la manosa es un carbohidrato que solo se encuentran en procariotas de forma que el leucocito puede saber así que no es una célula nucleada e iniciara una respuesta inmune.
33 los receptores de membrana de la inmunidad innata reconocen los patrones moleculares de familias de gérmenes y estos por lo general son carbohidratos. Por ejemplo la manosa es un carbohidrato que solo se encuentran en procariotas de forma que el leucocito puede saber así que no es una célula nucleada e iniciara una respuesta inmune.
35 de la inmunidad innata podemos decir:
-sistema muy conservado y evolucionada presente en plantas y animales
-se basa en una respuesta inflamatoria iniciada en macrófagos, polimorfonucleares y mastocitos.
- los defectos de este sistema son siempre letales
-reconocimientos de patógenos mediados por receptores (PAMP)


32 los receptores de la inmunidad
adaptativa son muy específicos para cada germen y aun más para cada molécula
del germen de forma que pueda reconocerlo por ese ligando especifico que tiene. En este tipo de inmunidad literalmente se puede generar un
receptor para cualquier cosa que este hecho de moléculas.
34 los receptores de la
inmunidad adaptativa son los receptores de las
células T y las inmunoglobulinas.

36 los patrones del
reconocimiento de la inmunidad:
En la inmunidad innata los
patrones pueden llegar a unas cuantas decenas porque la cantidad de
carbohidratos presentes en la membranas de los patógenos también es poca. Por
ejemplo si el elemento que esta el identificación tiene un receptor para RNA de
doble cadena entonces será catalogado como un patógeno porque solamente las
bacterias tiene RNA de doble cadena en los humanos es de una sola cadera.


37 en el caso de la
inmunidad adaptativa se pueden presentar cientos de millones de posibilidades
en el reconocimiento de una sola molécula de la membrana que distinga al
patógeno.
38 la inmunidad innata
está presente en todos los animales mientras que la inmunidad adaptativa
solamente se encuentra presente a partir de los peces.
39 uno de los receptores
de la inmunidad innata son los receptores tipo TOLL o TLR que van a reconocer
bacterias hongos y virus.

INMUNIDAD INNATA
FISIOLOGÍA DEL GLÓBULO BLANCO

INMUNIDAD INNATA
FISIOLOGÍA DEL GLÓBULO BLANCO
GRANULOCITOS
-Constituyen el mayor
porcentaje de los leucocitos en la sangre periférica
-Ejercen función de defensa
del organismo contra los procesos infecciosos
-Los granulocitos incluyen:
-neutrofilos
-eosinofilos
-basofilos
41 clasificación general
de los glóbulos blancos:
Por la presencia de gránulos:
-granulocitos
-agranulocitos (linfocitos
y monocitos)
Por su origen
-mieloide
-linfoide
Por las características
del núcleo:
-monomorfonucleares
-polimorfonucleares
Según su función:
-procesos fagociticos (
neutrófilos y monocitos)
-procesos inmunitarios (linfocitos)
42 los polimorfonucleares
se destacan porque son los más numerosos y los más activos.
NEUTROFILO
NEUTROFILO
43 como ya dijimos los
neutrófilos sufren un procesos de maduración(diferenciación) pasando de ser
mieloblastos a promielocito después a mielocito neutrófilico y a célula
neutrofilica en banda y finalmente a neutrófilo.

En este proceso de diferenciación se observa la progresiva condenzación y segmentación del núcleo con lo cual el nucleo se está inutilizando de forma parecida a lo que sucedía con el eritrocito solo que en este caso no se expulsa el núcleo. Esta proceso de ir desactivando al núcleo ocasiona que el neutrófilo no tenga mucho tiempo de vida.

En este proceso de diferenciación se observa la progresiva condenzación y segmentación del núcleo con lo cual el nucleo se está inutilizando de forma parecida a lo que sucedía con el eritrocito solo que en este caso no se expulsa el núcleo. Esta proceso de ir desactivando al núcleo ocasiona que el neutrófilo no tenga mucho tiempo de vida.
44 la forma joven del
neutrófilo es el neutrófilo abastonado porque se observa que recién se está
segmentando.


45 una infección en el
organismo causa una mayor producción de neutrófilos y más aun si esta infección
es bacteriana. Se elevan en gran medida los neutrófilos abastonados lo cual
indica que se están produciendo en mayor medida.
46 los neutrófilos son los
primeros elementos de la defensa en acudir a la zona de agresión y se da en
base a sustancias quimiotacticas que son sustancias que van a llamar a los
neutrófilos. Los factores derivados de la coagulación son quimiotacticos.


47 los abscesos se forman justamente por la reunión de neutrófilos en la zona de la herida.

48 la infección o
inflamación puede ser de dos tipos:
Aguda: comienzo rápido, duración corta, edema y predominio de neutrófilos
Crónica: progresiva,
duración larga, predominio de macrófagos y linfocitos, angiogénesis, fibrosis y
necrosis tisular.
49 por lo visto decimos
que si los neutrófilos ya no pueden controlar la infección entonces sobrevendrá
la inmunidad adaptativa. Por ejemplo la tuberculosis en franco proceso de desarrollo induce la formación de linfocitos, es decir de la inmunidad adaptativa. Tiene este comportamiento porque la tubersulosis es una infección crónica, de manera que, la inmunidad innata da paso a la inmunidad adaptativa.
50 los neutrófilos tienen
mecanismos enzimáticos y químicos para eliminar a las bacterias. Los
neutrófilos poseen:
-gránulos primarios.
Contiene a las mielo-peroxidasas que va a formar al ión hipocloroso.
-los gránulos secundarios
contiene elementos enzimáticos

MONOCITOS
51 los monocitos son
también células de la defensa o la inmunidad innata. Monocito es el nombre que
toma esta célula cuando se encuentra en el torrente sanguíneo. Cuando los
monocitos migran hacia otros tejidos se les denomina macrófagos tisulares.
52 los monocitos penetran
en la sangre periférica desde la medula ósea y circulan por aproximadamente 72
horas.
53 los monocitos pueden
vivir de dos a tres meses y nunca más regresan a la circulación.


Veremos que los monocitos no se comportan de la misma forma en los diferentes tejidos a donde emigran. estas diferencias dependen de morfología y fisiología del tejido en el que se desarrollan los monocitos.

54 lo normal es que haya de 5-10 mil leucocitos por milímetro cubico.
56 los macrófagos engullen a las bacterias y otros elementos extraños al organismo por medio de los fagosomas, que son vesículas que se forman en la membrana del macrófago y luego en el interior se unen a vesículas que contiene enzimas degradativas.


FORMULA LEUCOCITARIA


54 lo normal es que haya de 5-10 mil leucocitos por milímetro cubico.
56 los macrófagos engullen a las bacterias y otros elementos extraños al organismo por medio de los fagosomas, que son vesículas que se forman en la membrana del macrófago y luego en el interior se unen a vesículas que contiene enzimas degradativas.

MECANISMOS DE ELIMINACIÓN DE LOS GERMENES
Pueden ser de dos tipos:
-mecanismos dependientes del oxigeno:se general radicales libres que oxidan a la membrana del germen.
-mecanismos no dependientes del oxigeno:que se realizan a través de encimas.
-mecanismos dependientes del oxigeno:se general radicales libres que oxidan a la membrana del germen.
-mecanismos no dependientes del oxigeno:que se realizan a través de encimas.

FORMULA LEUCOCITARIA

VARIACIONES FISIOLÓGICAS
DE LA FORMULA LEUCOCITARIA
55 la cantidad de
leucocitos puede variar de manera normal de acuerdo a la edad
Recién nacido :10-25 mil
Un año de edad:6-18
mil
Edad pre-escolar: 6-15 mil
Edad escolar: 5-13 mil
Adulto: 5-10 mil
La cantidad de leucocitos
también varia con:
-el ritmo circadiano
-condición de reposo
corporal: tendencia a la leucopenia
-condición de ejercicio:
tendencia a la leucocitosis


LAS PROTEÍNAS DEL
COMPLEMENTO
Inmunidad innata
Inmunidad innata
57 el objetivo de las
proteínas del complemento es formar poros en la membrana del patógeno de forma
que este al tener mucho más solutos dentro de la membrana por difusión la
célula se llenara de agua hasta que estalla. Se busca la destrucción del patógeno por estallido.
58 complejo de ataque de
membrana: es el poro que se forma por las proteínas del complejo para hacer
estallar el patógeno. Estas proteínas se encuentran disueltas en el suelo de la sangre. La formación del complejo de ataque de membrana es un procesos similar a la cascada de la coagulación, por ello es razonable hablar de la cascada del complemento.


59 para que se forme el
complejo de ataque de membrana tiene que haber tres fracciones
importantes, que son los productos más
importantes del complemento:
-C3a: inflamación
-C3b: opsonozación y
fagocitosis
-C5a inflamación


60 existen tres formas de
activar la vía del complemento:
-La vía alterna: microbios
pueden activas el complemento
-la vía clásica: vía de
los anticuerpos
-la vía lectina : por la manosa(bacterias)
-la vía lectina : por la manosa(bacterias)
61 con un poco de más
detalle podemos decir que la vía de las lectinas y de los anticuerpos confluyen
para activar al C5 de manera que este se divide en C5a y C5b este último es el
que entrará en acción uniéndose a:C6, C7, C8, C9 de manera que así forman el
poro.



62 el objetivo de la
cascada del complemento es la formación de C5b.
63 C9 es realmente en que
forma el poro de la membrana atravesando completamente a la membrana lo demás
era solamente un marcador de membrana

VÍA CLÁSICA
64 en la via clásica el
antígeno-anticuerpo activan al C-1 y este va activar al C-4 y C-2 a partir de
esto se forman C4a y C4b también se forma C2a y C2b de suerte que su unen C4b
con el C2a y tenemos C4b2a que se unen a la membrana. Este complejo se denomina
convertasa de C3 de forma que dividen al C3 en C3a y C3b. el C3b se une al
complejo constituyendo: C4b2a3b , este nuevo complejo se denomina
convertasa de C5 de forma que activan al C5 convirtiéndolo en C5a y C5b
entonces C5b que es la más importante se unirá como ya dijimos a C6,C7, C8 y C9
para formar el complejo de ataque formando el poro.


Observemos que C1 se une al anticuerpo-antígeno. De una forma especial vemos que debe estar unida a dos anticuerpos que se encuentran unidos al antígeno es decir están en una estrecha relación con los anticuerpos y los antígenos de manera que así se da la activación de C1.



Observemos que C1 se une al anticuerpo-antígeno. De una forma especial vemos que debe estar unida a dos anticuerpos que se encuentran unidos al antígeno es decir están en una estrecha relación con los anticuerpos y los antígenos de manera que así se da la activación de C1.

VÍA ALTERNATIVA
65 la vía clásica es la primera vía que se descubrió pero no es la principal. A vía alterna si es la principal.
66 la vía alterna actúa sobre
la superficie de las bacterias que es donde se encuentran diversos
polisacáridos. Por ello podemos decir que esto forma parte de la inmunidad
innata.
67 la superficie
bacteriana cliva al C3 en C3a y C3b gracias al factor B y P se forma C3bBbP que
es la convertasa de C3 por lo cual se forma C3a yC3b de manera que este ultimo
complemento se une a la convertasa previamente descrita formando C3bBb3bP la
cual es la convertasa de C5 después la historia es la misma.


68 por lo que vemos los factores del complemento
van desde el C1 hasta el C9. Son proteínas que están sueltas en el plasma.
69 los factores del complemento hacen un trabajo
que forma parte de la inmunidad innata reconociendo a las bacterias que entran
en el organismo. todo este proceso es muy importante porque nos permite
sobrevivir en un medio plagado de bacterias.
VÍA DE LAS LECTINAS
VÍA DE LAS LECTINAS
70 la vía de las lectinas
utiliza una molécula que es el MASP que actua sobre C4y C2 de forma que se
forma luego de su activación C4b2a que es la misma convertasa de C3 de la vía
clásica siguiendo así la misma vía. Esta vía utiliza la manosa de las bacterias para su activación.


71 Los
elementos residuales son el C3a y el C5a son muy tóxicos cuando se
encuentran libres. Estas son conocidas como anafilotoxinas que causan inflamación y una fuerte reacción anafilactoide en el organismo. Recordemos que la reacción anafiláctica es causada por la inmunoglobulina E, se trata de una reacción alérgica donde se libera fundamentalmente histamina y se produce el edema. C3a y C5a causan una dilatación de los vasos sanguíneos produciendo un edema
generalizado, que a su vez puede llevar al individuo a un shock y posiblemente la muerte. Como vemos la reacción anafilactica o alergica se parece mucho a la reacción anafilactoide.




72 las proteínas del
complemento son activadas también por la inmunoglobulina M que es muy
fuerte a diferencia de la inmunoglobulina G que no es tan potente.Un caso interesante ocurre
cuando se produce una reacción anafilactoide mediada por IGM y activada por C3a y C5a a una persona cuyo tipo de sangre es A y le hacen una
transfusión con sangre de tipo B (incompatibilidad ABO) por error. Los anticuerpos de la sangre transfundida atacan a los glóbulos rojos del paciente a los cuales toma como antígenos, produciendose inmediatamente la hemolisis. Ahora recordemos que los
anticuerpos A y B son inmunoglobulinas M (que pueden generar C3a y C5a) a diferencia de el factor Rh que es
una inmunoglobulina de tipo G. Por este motivo la reacción ante el antígeno es muy fuerte y puede causar un shok
inmediatamente. Efectivamente se produce una reacción
anafilactoide por la liberación de C3a y C5a.
73 el C3b también tiene otra función a parte de formar la
convertasa de C5 que es ser opsonil(marcador) para el macrófago. El macrófago tiene
receptores para el C3b y esta última tiene la capacidad de unirse a la membrana
de la bacteria de manera que sirve como un elemento de unión entre la bacteria
y el fagocito para que se de el proceso de fagocitosis. Podemos decir que es
una forma de marcar la basteria con C3b


INMUNIDAD ADAPTATIVA
74 debemos decir que la
inmunidad innata es rápida pero inflexible porque una vez que surge un nuevo patógeno por
evolución este no tiene como reconocerlo.
75 la síntesis de nuevos
receptores para el reconocimiento de nuevos antígenos se denomina inmunidad adquirida.
76 los nuevos receptores son producidos por los linfocitos T (receptores t) y los linfocitos B con la
producción de anticuerpos.
77 estos receptores potencialmente pueden
ser modificados hasta en 100 mil millones de variedades diferentes.

En la figura de arriba observamos como el ingreso de un patógeno en principio no puede ser neutralizado debido a que el macro fago no presenta los receptores para este tipo de patógeno. Después de algún tiempo los linfocitos son capaces de producir los receptores adecuados en las inmediaciones de sus membranas celulares. estos receptores son liberados en el plasma para que puedan ser acoplados a los macrófagos. De esta forma los macrófagos tiene el receptor especifico para el nuevo patógeno y pueden empezar a destruirlos.

En la figura de arriba observamos como el ingreso de un patógeno en principio no puede ser neutralizado debido a que el macro fago no presenta los receptores para este tipo de patógeno. Después de algún tiempo los linfocitos son capaces de producir los receptores adecuados en las inmediaciones de sus membranas celulares. estos receptores son liberados en el plasma para que puedan ser acoplados a los macrófagos. De esta forma los macrófagos tiene el receptor especifico para el nuevo patógeno y pueden empezar a destruirlos.
INMUNIDAD HUMORAL
78 la inmunidad humoral se encuentra en el plasma. Y es producida por los anticuerpos.
79 el anticuerpo esquemáticamente tiene la forma de una "Y" donde observaremos una zona variable(uno de los brazos), es decir de diferente constitución molecular, la cual se une al antígeno. Esta zona también es llamada región FAB (región de anticuerpo). También tiene una región constante (el pie de la Y) presente
en todos los anticuerpos. esta región también se le denomina región FC (región cristalizable).


En la imagen de abajo observamos como los anticuerpos disponen de una excelente flexibilidad para uninierse al antígeno correspondiente:
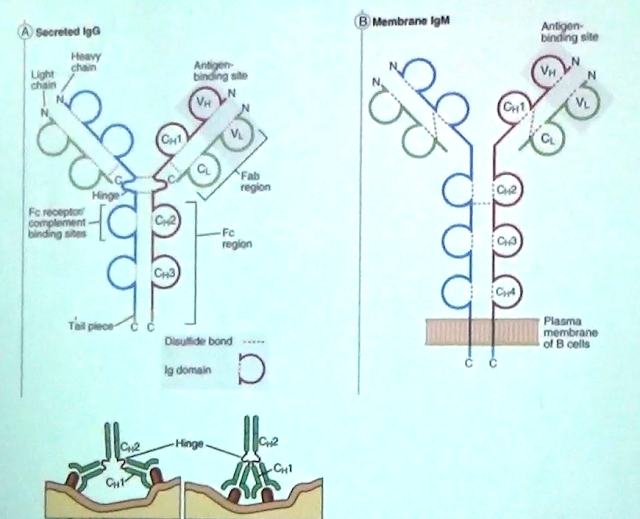
80 el anticuerpo y el antígeno se atraen por diferentes fuerzas:

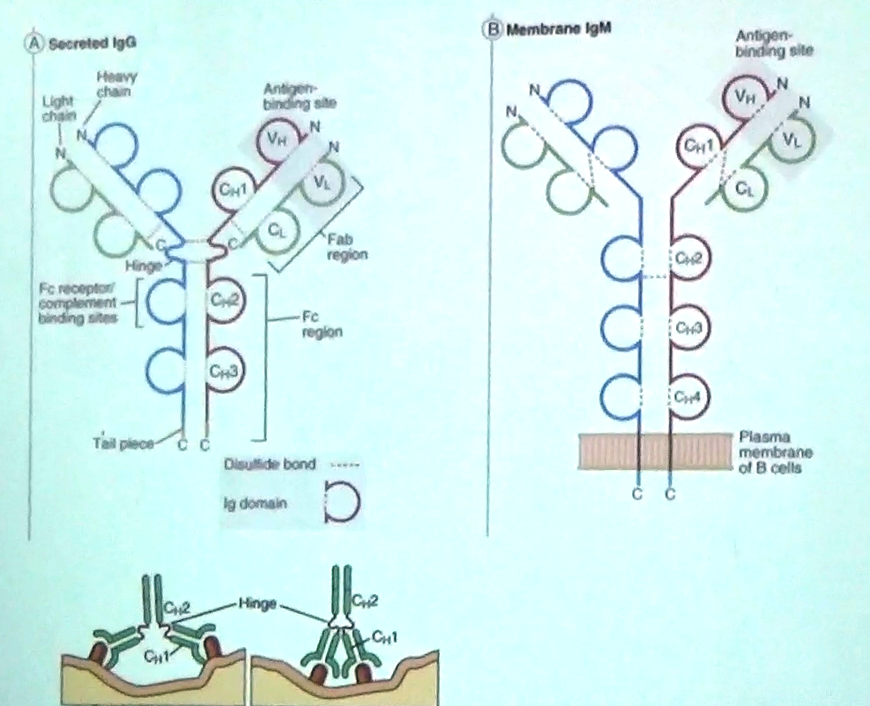
80 el anticuerpo y el antígeno se atraen por diferentes fuerzas:

81 los anticuerpos son de
5 tipos. El prototipo de las inmunoglobulinas es la inmunoglobulina G que
parece una “Y”.
82 la inmunoglobulina A
que son dos “Y” unidas por un péptido.
83 la inmunoglonulina M es
un pentámero.
84 La
inmunoglobulina M es mucho mas efectiva que la inmunoglobulina G porque esta
ultima solamente tiene dos sitios de unión mientras que la inmunoglubulina M
tiene 10 sitios de unión.


85 la inmunoglobulina M se
conoce como aglutinina.
86 una explicación de
porque en la prueba de reconocimiento del grupo sanguíneo en el sistema ABO la
aglutinación (formación de grumos) es muy marcada es por la acción inmunoglublima M, mientras que en la prueba del reconocimiento del factor Rh (sistema Rh) actúa la
inmunoglobulina G. Este tipo de diferencias en la aglutinación queda muy bien explicada cuando pensamos en que la inmunoglobulima M posee 10 sitios de únión mientras que la inmunoglobulima G solamente posee 2. El poder de aglutinación de la Ig. M es mucho mayor que la de la Ig. G
87 en la inmunoglobulina G
encontramos una cadena pesada y una cadena ligera. precisamente lo que
diferencia a los anticuerpos es la cadena pesada. En la imagen de abajo se representa a la cadena pesada en rojo, mientras que la cadena ligera se representa en azul:


88 los encargados de
producir los anticuerpos son los linfocitos B
89 los linfocitos B cuando
están preparados para producir anticuerpos se convierten en células plasmáticas
(plasmocitos)
90 por ello diremos que
los linfocitos B son las células más basofilas del organismo porque producen
una alta cantidad de proteínas.
91 la célula madura
funcional del linfocito B es la célula plasmática.

92 las funciones de las inmunoglobulisnas:
NEUTRALIZAR: cuando una bacteria ingresa al ornanismo los anticuerpos se adieren a la bacteria de forma que impiden que la bacteria se una a los tejidos del organismo. La opsonización tambíen se puede presentar frente al ingreso de toxinas al organismo.




OPSONIZACIÓN: si el patógeno ingresa al organismo inmediatamente los anticuerpos se adhieren a ciertas moléculas que se encuentran en la membrana de del patógeno. Es notable la diferencia de tamaño que se presenta entre el anticuerpo y el patógeno como por ejemplo una bacteria. Las diferencias de tamaño impiden que el anticuerpo pueda aglutinar a la bacteria y producir su eliminación por completo. La función del anticuerpo es "marcar" a la bacteria de forma que los macrofagos puedan unirse a los patógenos a través de de los anticuerpos. Recordemos que los anticuerpos tienen una región FC, que constituye la región de unión para el macrofago a través del receptor para FC.



En la imagen de arriba observamos como el anticuerpo funciona como un puente entre la bacteria y el macrofago. además del anticuerpo se observa la presencia de C3b que
también actúa como un opsonozador. Por lo tanto las opsoninas serían:
*inmunoglobulima G
*C3b
la inmunoglobulima M se caracteriza por se aglutinante y activador del complemento.




En la imagen de arriba observamos como el anticuerpo funciona como un puente entre la bacteria y el macrofago. además del anticuerpo se observa la presencia de C3b que
también actúa como un opsonozador. Por lo tanto las opsoninas serían:
*inmunoglobulima G
*C3b
la inmunoglobulima M se caracteriza por se aglutinante y activador del complemento.

En la figura que presentamos arriba se observa como se pueden presentar las diversas formas de opsonizar a los patógenos tanto mediante un anticuerpo como por el C3b. Es notable el resultado que produce opsononizar una bacteria mediante la inmunoglubulina G y el C3b
-activación del complemento
-activación del complemento
-aglutinar: por la IgG.

DIFERENCIAS ENTRE LAS INMUNOGLOBULINAS
Para activar el complemento solamente necesitamos una IgM o bien dos Ig G. Esto se debe a que la IgM tiene 10 brazos, mientras que la IgG tiene solamente dos brazos. La IgG es muy efectiva para opsonizar pero no para aglutinar ya que debido al potencial Z dos células no se pueden tocar. Las membranas de células idénticas tienen la misma carga eléctrica de este modo se presenta una fuerte repulsión entre ellas. la IgG tiene brazos cortos y no puede unir dos células por el corto espacio que quedarian entre estas con una IgG.

Otra diferencia se presenta en cuanto a la localización de la Ig. por ejemplo la Ig. se encuentra en las secreciones. la IgG presenta las mayores concentraciones y mayor vida media en suero. aunque la de mayor producción es la IgA no se concentra porque su recambio es continuo.

La inmunoglobulina tipo G presenta cuatro tipos. solamente las den tipo uno y tres pueden atravesar la barrera placentaria. cuandoe sto ocurre puede existir la posibilidad de presentarse la eritoblastosis fetal o enfermedad hemolitica del recien nacido cuando la madre es Rh- y el feto es Rh+.
INMUNOGLOBULINA A

INMUNOGLOBULINA M

Otras inmunoglobulinas:

gh
PRACTICA DE
FISIOLOGÍA DE LA SANGRE
TEORIA EN PIZARRA
1 El proceso de la
hemostasia es aquel por el cual la sangre se mantiene en su funcionamiento
normal de manera que este fluido cumple con sus funciones adecuadamente.
2 la hemostasia se puede
dividir en cuatro etapas:
-primero:
vasoconstricción, esta solamente se da en los vasos de mediano calibre porque
la presión es demasiado fuerte en los vasos de gran calibre mientras que el los
capilares solamente se puede dar la regulación del flujo. Para la vasoconstricción
existen dos mecanismos
Neuronal: el sistema simpático produce
la vasoconstricción
Endotelial: la secreción
de endotelina por parte del endotelio que también produce el efecto de la
vasoconstricción.
-segundo: agregación
plaquetaria- la unión de las plaquetas a la membrana celular se da por el
factor de von willebrand. Este fenómeno propicio la adhesión la secreción y la
agregación de las plaquetas. La adhesión se da gracias a las selectinas y la
secreción quiere decir que las plaquetas van a liberar los gránulos que estas
contiene es decir los gránulos alfa(selectina, fibrinógeno, factor V, VIII) y
los gránulos delta(ADP, ATP, calcio, histamina y serotonina). En cuanto a la
agregación sabemos que se da entre plaquetas.
-tercero: cascada de la
coagulación o hemostasia secundaria. Donde se forma un tapón plaquetario
inicial.
-cuarto: formación del
tapón plaquetario en si. Que presenta fibrina polimerizada. Con una activador
el plastinógeno tisular, esta es una molécula que funciona como anticoagulante
y sirve para mantener al coagulo circunscrito solamente a un área y que este no
se extienda demasiado.
3 es muy importante saber
que existen dos vías de la coagulación:
-vía intrínseca: esta vía
se inicia con el factor número XII o factor de Hagen.
-vía extrínseca: esta vía
se inicia con la estimulación exógeno y por la participación del factor tisular
4 debemos saber que el
factor XII, es decir inicia la cascada de la coagulación.
5 para medir la vía
intrínseca se usa el tiempo de tromboplastina(factor XI) parcial: TPT.
6 para medir la vía
extrínseca se una el tiempo de protrombina (factor II): TP
7 las dos pruebas
mencionadas analizan los factores V,II, y X que pertenecen a la vía común.
Mientras que TP analiza también de manera exclusiva el en factor VII y la
prueba de TPT analiza exclusivamente los factores: XII,XI,IX VIIIy I.
8 para realizar la TPT lo
que se hace es utilizar partículas de carga negativa. Mientras que para hacer
la prueba de TP se hace añadiendo fosfolípidos tisulares.
9 el endotelio vive en un
equilibrio entre elementos pro-tromboticos y anti-tromboticos. Esto se da
debido a que es a la hemostasia es un proceso fisiológico es decir muy
necesario.
10 dentro de los elementos
pro-tromboticos podemos ligados a las plaquetas y a la membrana del endotelio
podemos considerar al factor de von willebrand.tambien el factor tisular es un
elemento pro-trombotico.en el plasma podemos encontrar al inhibidor del
activador de plastinógeno.
11 dentro de los elementos
antitromboticos es decir anticoagulantes podemos tener la heparina en relación
a las plaquetas y en el plasma el activador de plastinogeno
12 en cuanto a la cascada
de coagulación como ya dijimos son dos vías:
Vía extrínseca: se inicia
con un factor externo( destrucción de células de la piel) que liberan el factor
tisular este se une al factor VII en un fosfolípido de la membrana de la
plaqueta mediado por el calcio y se forma un complejo capaz de activar al
factor X.
Vía intrínseca: en esta
vía se dan procesos más complejos. diremos así que el factor XII se activa
iniciando la cascada de la coagulación. Una vez activado este factor activa al
factor XI (tromboplastina)con ayuda del factor II activado ( trombina). Una vez
activado el factor XI va activar al factor IX(enzima) este se va a unir al VIII
(cofactor) activado de manera que junto al calcio y unidos a un fosfolípido de
la membrana forman un complejo de proteico. Este complejo es capaz también de
activar al factor X.
13 como vemos las dos vís
sirven para activar al factor X de maneta que este factor activado se une al
factor V(cofactor y se necesita la intervención del factor II) que junto a factor plaquetario III y por
mediación del calcio formaran un complejo proteico enzimático denominado
complejo protrombinasa.
14 seguidamente el
complejo protrombinasa hace posible la activación de la protrombina en trombina
y la trombina es decir el factor IIa activa al factor I y lo convierte en
factor Ia es decir fibrina.
15 no olvidemos que la
fibrina para estabilizarse requiere del factor XIIIa
16 debemos apuntar que la
vía extrínseca es más rápida y no requiere de trombina para su acción sobre el
factor X.
17 no olvidar que el
factor XII activa también otras tres cascadas
MESA DE PRACTICA
-el VCG es la prueba por
la cual podemos saber en cuento tiempo los elementos formes de la sangre se
separan del plasma. esta prueba nos da una idea de la cantidad de proteínas que
pueda hacer en la sangre.
-recordemos que la
hemoglobina y el hematocrito está en la proporción de uno a tres porque la
cantidad de hemoglobina que ocupa en un eritrocito es la tercera parte de este
eritrocito. Por ejemplo si tenemos 15 de hemoglobina tendremos 45 de
hematocrito.
-
-El macro-hematocrito es
más preciso que el micro-hematocrito. Pero dura más en total media hora en
cambio en micro-hematocrito dura solamente cinco minutos.
-cual es la relación
hemoglobina- hematocrito es de 1 a 3
-en la membrana de los
eritrocitos hay ácido cialico el cual le brinda al eritrocito una carga
negativa a los eritrocitos. De forma que las membranas se repelen, pero se da
el aumento de proteínas en la sangre debido a una inflamación por ejemplo
entonces estas tienden ubicarse entre las membranas de manera que redicen la
repulsión por lo cual las plaquetas tienden a sedimentarse por la fuerza de la
gravedad. Es decir la velocidad de sedimentación globular aumenta. Esta
velocidad es normal en hombres de 0-20 ml por hora en hombres de y de 0-15 ml
en mujeres por hora.
- a grandes rasgos existe
la coagulación primaria y la coagulación secundaria o la homeostasis primaria y
la homeostasis secundaria.
-la hemostasia primaria
comprende la vasoconstricción y la acción de las plaquetas. Es decir la fase
vascular y la fase plaquetaria.
-la hemostasia secundaria
consiste en la formación del coagulo secundario en base a la polimerización del
fibrinógeno y se convierte a fibrina.
- la prueba para ver si
existe algún problema en la hemostasia primeria es la priva de la sangría.
Donde provoca una pequeña herida de forma que vamos registrando con el
cronometro el tiempo e que tarda en dejar de sangrar la herida normalmente este
tiempo de es de 0 a 2 minutos. PRUEBA DE TIEMPO DE SANGRIA
-si queremos evaluar la
fase de la coagulación es decir la hemostasia secundaria o coagulación
secundaria entonces hacemos la prueba del TIEMPO DE COAGULACION
-se usaron los anticuerpos
Antia A: si reacciona con
este anticuerpo entonces el paciente es del grupo A, se observa la formación de
coágulos
anti B: si reacciona con
este anticuerpo entonces el paciente es del grupo B, se observa la formación de
coágulos
anti D: si reacciona con
este anticuerpo quiere decir que es rh+ y si no rh-
-si reacciona con los
anticuarpos A y B entonces es del grupo AB
-el tiempo de coagulación
es más exactos cuando se hacen varias
pruebas y se saca el promedio.
RESUMEN DE LA GUÍA PRACTICA
PRACTICA 1
DETERMINACIÓN DE LA CONCENTRACIÓN DE HEMOGLOBINA. HEMATOCRITO. CONSTANTES
CORPUSCULARES. VELOCIDAD DE SEDIMENTACIÓN GLOBULAR.
1 Buscamos la determinación relativa
de los elementos que constituyen la sangre.
2 Obtenidos los datos de la hemoglobina, hematocrito y numeración de
hematíes, se puede calcular el volumen de cada uno de los hematíes y la
cantidad absoluta y porcentual de hemoglobina por glóbulo rojo. Índices que son
de suma utilidad para la interpretación de las alteraciones de los hematíes,
cuando las hay, y para deducir, en muchos casos el origen de dichas alteraciones.
DETERMINACIÓN DE LA CONCENTRACIÓN DE HEMOGLOBINA

HEMATOCRITO
3 el hematocrito es la relación porcentual entre la cantidad de elementos
formes y el plasma o en forma más restringida el porcentaje del volumen ocupado
por eritrocitos con respecto al volumen total de sangre.
4macrohematocrito (wintrobe)
Se utiliza heparina
Centrifugar por 30 minutos a 3000 revoluciones por minuto.
5 microhematocrito:
Se usa el tubo capilar
Se centrifuga por 5 minutos a 10000 RPM

CONSTANTES CORPUSCULARES
6 volumen corpuscular medio:
volumen que por término medio tiene un hematíe
7 el VCM esta arriba de 97 en las anemias macroliticas, y a menor de 83 en
las anemias microliticas.


8 hemoglobina corpuscular media:
indica la cantidad de hemoglobina promedio que tiene un hematíe.

9 se habla de hipercromía cuando el HCM es mayor que 31 (en caso de anemia
permisiosa o megaloblastica) de otro lado de habla de hipocromía cuando los
valores de HCM están por debajo de 27 (anemia ferropénica)
10 concentración de hemoglobina
corpuscular media. Contenido medio de hemoglobina por unidad de volumen
eritrocitarioo hematocrito.

11 valores inferiores al normal se observan en las anemias hipocromicas
VELOCIDAD DE SEGMENTACIÓN GLOBULAR (VSG)
12 en una muestra anti-coagulada depende de la tendencia a aglutinarse que
tienen los eritrocitos.
Fuerzas de atracción: van der waals
Fuerzas de repulsión: fuerzas Z algunos factores disminuyen esta fuerza
tales como las globulinas el firinogeno la albumina no lo hace
13 si las fuerzas de repulsión se ven disminuidas entonces se aglutinará
mas rápido ello quiere decir que hay un aumento de proteínas por ejemplo debido
a una infección.las hiperproteimias aumentan la viscosidad plasmática
aumentando el VSG
14 en esta prueba se una en oxalato un anticoagulante

PRACTICA 2
FRAGILIDAD OSMOTICA. LA HEMOSTASIA: TIEMPO DE SANGRIA, COAGULACIÓN Y
RECALCIFICACIÓN.
FRAGILIDAD OSMOTICA.
1 cuando la destrucción de eritrocitos es muy acelerada causa anemia
2 la hemolisis excesiva depende de la mayor fragilidad de los eritrocitos o
una resistencia disminuida (esferocitosis hereditaria, anemia hemolítica
congénita)
3 el aumento de la fragilidad esta determinada por dos factores:
-es inveramente proporcional a la concentración de hematíes
-es directamente proporcional al grosor de la membrana del hematíe.
4una fragilidad osmótica disminuida o una resistencia aumentada denota un
aplanamiento excesico del hematíe.
5 la concentración máxima de cloruro de sodio con una hemolisis minima
determina la hemolisis inicial; la concentración mas alta con la que la
hemolisis sea completa expresa la hemolisis total.
6 una hemolisis incipiente con concentraciones mayores de
cloruro de sodio es señal de que la fragilidad osmótica de los hematíes ha
aumentado. La fragilidad osmótica de los hematíes esta aumentada si hay
hemolisis a concentraciones de más de 0.5 % de cloruro de sodio. Por otro lado
la fragilidad osmótica esta disminuida si la hemolisis es incompleta en cloruro
de sodio al o.3%
TIEMPO DE SANGRIA
1 mide la duración de la hemorragia, es decir mide el
tiempo en que se formo el primer tromboplaquetario.
2 mide la contractibilidad de los vasos sanguneos y la
capacidad de las plaquetas.
3 los valores normales de tiempo de angria son de 1 a 5
minutos.
4 este tiempo se encuentra alargado en las trombopenias.
TIEMPO DE COAGULACIÓN
1coagulacion in vitro
2 estudia la coagulación intrínseca es decir la tercera
fase de la hemostasia.
3 no se una anticoagulante
4 el valor normal de tiempo de coagulación es de 4-10
minutos.
5 se encuentra alargado en la carencia de factores
plasmáticos de la coagulación, en las hipoprotrombinemis por falta de vitamina
K o insuficiencia hepática
TIEMPO DE RECALCIFICACIÓN
1 consiste en medir el tiempo de coagulación del plasma,
separado al hacer la sangre incoagulable mediante citrato y añadiéndole
nuevamente calcio.
2 al agregar el cloruro de calcio al suero no coagulado
se ve la formación de una película de fibrina.
3 los valores normales de recalcificación son de 70-170
segundos.
PRACTICA 3
DETERMINACIÓN DEL GRUPO SANGUINEO
1en la membrana de los hematíes están los antígenos (ABO)
y Rhesus
2 en el plama existen anticuerpos: antiA y antiB estos
anticuerpos son inmunoglulinas M de tipo aglutinante que no atraviesan la
placenta
3 todos los anticuerpos anti-D son de tipo adquirido y
atraviesan la placenta.
INCOMPATIBILIDAD DEL GRUPO SANGUINEO
1 en el sistema ABO una incompatibilidad es muy grave en
un error de transfisión.
2 esta reacción es iniciada por la unión
antígeno-anticuerpo
3 la reacción de incompatibilidad ABO es prototipo de las
reacciones mediadas por anticuerpos de tipo IgM, diferente de las reacciones mediadas
por IgG
4 el tipo sanguíneo A tiene el antígeno A y en el plasma
el anticuerpo B
5 el tipo sanguíneo B tienen el antígeno(aglutinógenos) B
y en el plasma en anticuerpo A (aglutina)
6 el tipo sanguíneo AB tiene antígeno AB y en el plasma
no tiene anticuerpos
7 el tipo sanguíneo O no tiene anticuerpos y en el plasma
tiene anticuerpos A Y B
8 en una incompatibilidad sanguínea lo que ocurre es que
los anticuerpos (aglutinas) se unen a varios eritrocitos a la vez, provocando
la aglutinación
TIEMPO DE PROTROMBINA TP
1 evalúa los factores de la vía extrínseca (VII, X,
V,II y fibrinógeno)
2
TIEMPO DE TROMBOPLASTINA TOTAL ACTIVADO (TTPA)
1 evalúa los factores de la vía intrínseca y común
(XII,XI,IX, VIII, X, V, II y fibrinógeno)
No hay comentarios:
Publicar un comentario